Среди вопросовъ частной систематики, издавна привлекавшихъ на себя вниманіе орнитологовъ, быть можетъ, ни одинъ другой не вызывалъ столь многочисленныхъ и столь же безуспѣшныхъ попытокъ разрѣшенія, какъ вопросъ о происхожденіи и предѣлахъ обширной группы птицъ, по численности обнимающей собою половину всѣхъ нынѣ извѣстныхъ видовъ, а по составу — формы съ наивысшимъ типомъ развитія организаціи даннаго класса. Mutatis mutandis, можно было бы сказать, что «отрядъ» Passeriformes въ отношеніи всѣхъ прочихъ птицъ напоминаетъ изолированное положеніе послѣднихъ въ ряду позвоночныхъ: то же обиліе видовъ, то же сходство и однообразіе общаго строенія, то же богатство переходныхъ формъ и частныхъ уклоненій на фонѣ чрезвычайно замкнутой въ себѣ организаціи. Прибавимъ сюда отсутствіе инструктивныхъ данныхъ палеонтологіи и крайнюю противорѣчивость фактовъ зоогеографіи, и намъ становится понятной исключительная трудность вышеуказанной проблемы. Любопытна исторія вопроса. Съ тѣхъ поръ какъ Ж. Кювье, руководимый геніальной интуи-ціей, впервые установилъ свой «Ordre Passereaux» въ объемѣ, близкомъ современнымъ Passeres, не преминувъ отмѣтить отрицательный характеръ ихъ отличій [7] , а L'Herminnier (1827), одинъ изъ наиболѣе проницательныхъ изслѣдователей того времени, остеологически обосновалъ воззрѣнія Кювье и въ первый разъ опредѣленно указалъ на характерное свойство этой группы — ея обособленность и однородность, послѣ того какъ Nitzsch, многосторонний и точный наблюдатель, по даннымъ каротидъ (1829) и птерилоза (1840) еще яснѣе очертилъ границы «Passerinae» и выключилъ изъ нихъ оставшуюся примѣсь инородныхъ формъ, объединенныхъ съ многими другими «лазащими» (Grimpeurs, Сuу.) во вновь установленный отдѣлъ Рісаrіае, — большинство послѣдующихъ авторовъ, игнорируя системы Nitzsch и L'Herminnier и не довольствуясь признаніемъ творца «Règne animal», во многомь уклонились отъ пути своихъ выдающихся предшественниковъ, стремясь найти во что бы то ни стало конкретный классификаціонный признакъ Passeres. Предпринятые съ этой цѣлью поиски по большей части оказались мало продуктивными (системы Reichenbach'a, Gray и Bonaparte), а нѣкоторыя удачныя открытія вначалѣ оказались роковыми. Таковы попытки Gloger'a (1834), охарактиризовавшаго всѣхъ «пѣвчихъ» Passeres — Aves Passerinao melodusae — длинною задняго ножного когтя и сложною мускулатурой Syrinx (въ отличіе отъ «Aves passerinae anomalae» = Picariae), таковы же указанія Cabanis'a на различія въ размѣрахъ и числѣ первостепенныхъ маховыхъ и тщательная разработка болѣе раннихъ наблюденій Kaiserling & Blasius о нѣкоторыхъ особенностяхъ плюсневыхъ покрововъ (установленіе 4 отдѣловъ Incessores со включеніемъ Picariae Питча), таковы, наконецъ, классическія изслѣдованія І. Мюллера надъ измѣненіями голосового аппарата Passeres, поведшія къ раздѣленію послѣднихъ на Oscines (Polymyodi), Tracheophonae и Picarii. Повторяемъ, эти немногіе вновь добытые признаки были куплены дорогой цѣной — вторичнымъ смѣшеніемъ «дятловыхъ» и «воробьиныхъ», и въ этомъ отношеніи Gloger, Cabanis и Müller съ внѣшней стороны вернулись болѣе чѣмъ за 100 лѣтъ назадъ, къ Линнееву порядку Рісае, группѣ чрезвычайно сборной и искусственной, а послѣдствія этой ошибки надолго отразились и въ позднѣйшей орнитологической литературѣ, въ формѣ продолжавшагося сближенія двухъ совершенно разнородныхъ группъ (системы Eyton, Gervais, Sundevall и многія другія). Что касается послѣдующихъ работъ, то положительные результаты ихъ сказались главнымъ образомъ на систематикѣ самихъ Passeriformes, на принципахъ классификаціи внутри самой этой группы и вовсе не коснулись собственно интересующаго насъ здѣсь вопроса — объ ея характеристикѣ in toto. Достаточно напомнить, что даже Gloger'oвскій признакъ — длина задняго когтя, — особенность и понынѣ приводимая во многихъ опредѣлителяхъ и считавшаяся единственной діагностичной, въ дѣйствительности не оказалась таковою (Upupa!). Отсюда понятно то значеніе, которое издавна придавали той или иной попыткѣ найти въ организаціи Passeriformes признакъ діагностично-характерный для всего отряда, — значеніе, тѣмъ бо́льшее, когда въ 1868 году авторомъ новой такой попытки оказался величайшій зоологъ того времени. Казалось, что на этотъ разъ была разрѣшена задача, что Гексли удалось вполнѣ использовать анатомическіе признаки, неоднократно выдвигавшіеся и прежними изслѣдователями, но лишь попутно (Nitzsch), или въ поверхностной и догматичной формѣ (Cornay, 1847), что все несмѣтное количество видовъ съ ихъ безконечнымъ разнообразіемъ въ habitus’ѣ и столь же изумительнымъ количествомъ тончайшихъ переходовъ, съ ихъ общепризнаннымъ единствомъ внутренней организаціи, на практикѣ казавшимся неуловимымъ, что всѣ шесть тысячъ съ лишнимъ Passeres объединяются одной чертой строенія, фактически легко и просто находимой: къ общей наглядности остелогическаго критерія присоединилось то, что вслѣдствіе топографическихъ особенностей вновь найденнаго признака — комплекса нѣкоторыхъ лицевыхъ костей, нерѣдко остающихся, т.-е. уцѣлѣвающихъ въ сухой, отпрепарированной шкуркѣ — при отсутствіи скелета рѣдкой формы, — не исключалась возможность пользованія матеріаломъ, односторонне собраннымъ господствовавшей въ то время школой фаунистовъ- систематиковъ. Съ другой стороны, эта рельефность классификаціоннаго признака не мало облегчала его критику. Послѣдняя не замедлила явиться, и въ историческомъ введеніи къ настоящей работѣ, въ первой главѣ нашей статьи, мы ознакомились уже съ общей критической литературой по вопросу о фактическихъ основахъ Гексліевой системы и, въ частности, съ главнѣйшими поправками и возраженіями, касавшимися послѣдняго отдѣла «Оn the Classification of Birds», отряда эгитогнатовъ. Мы видѣли, что изъ обѣихъ группъ, его составляющихъ, подотрядъ «Cypselomorphae» вскорѣ былъ кассированъ выключеніемъ изъ названнаго типа Trochilus и Caprimulgus, какъ не удовлетворяющихъ вполнѣ Гексліеву опредѣленію эгитогнатизма (заостренный спереди сошникъ колибри, отсутствіе передняго сошниковаго развилка у козодоя), что среди членовъ второго подотряда, именно «Coracomorphae», найдены были отдѣльныя уклоненія въ строеніи названнаго элемента (напр., округло- притупленная вершина Vomer у Calyptomene), что далѣе въ весьма разнообразныхъ и систематически весьма удаленныхъ группахъ — за предѣлами Passeres — наблюдались формы съ различными чертами «эгитогнатизма» (примѣры: расширеніе передняго сошниковаго конца у Tetrao urogallus, тупой и вильчатый сошникъ у нѣкоторыхъ Limicolae, глубоко расщепленная вершина того же элемента у Megalaema), и что въ глазахъ позднѣйшихъ систематиковъ руководящій признакъ типа, — въ томъ смыслѣ какъ онъ понимался Гексли, — конфигурація вершины сошника — утратилъ постепенно свой діагностическій характеръ. Мы видѣли, что на ряду съ такою негативною оцѣнкой первоначальнаго критерія отдѣльные ученые, желая удержать самый принципъ классификаціи, стараются придать ему другое толкованіе, надѣются путемъ различныхъ коррективовъ сохранить его таксономическое значеніе. Отсюда новыя опредѣленія эгитогнатизма, послѣдовательно вводимыя нѣкоторыми авторами: предполагаемыя особенности гистологической закладки сошника (Паркеръ), его первоначальная парность (Паркеръ, Фюрбрингеръ), расположеніе смежныхъ элементовъ (Гарродъ). Въ первой главѣ настоящей работы, въ концѣ критическаго обзора литературы, мы подробно остановились на нѣкоторыхъ изъ этихъ толкованій, имѣвшихъ цѣлью раскрыть неуловимое дотолѣ нѣчто, дѣйствительно лежащее въ основѣ разбираемой структуры, и тогда же мы установили недостаточность этихъ добавочныхъ опредѣленій признака, неминуемо поведшихъ къ затемнѣнію вопроса объ его систематическомъ достоинствѣ и, наконецъ, къ постепенному умаленію такового. Въ слѣдующей затѣмъ второй главѣ мы на исторіи развитія лицевого черепа у Corvus frugilegus попытались выяснить существенныя свойства эгитогнатизма и, въ частности, морфологическую природу сошника, — элемента, спорность толкованія котораго и обусловила рядъ вышеприведенныхъ разногласій по вопросу о границахъ разбираемаго типа. Мы убѣдились, что сошникъ эгитогнатовъ, казавшійся на первый взглядъ простымъ и столь легко опредѣлимымъ, въ дѣйствительности представляетъ сложный комплексъ двухъ независимыхъ образованій, а именно, частей покровнаго скелета + нѣкоторые отдѣлы носового лабиринта. Выясненіемъ морфологической особенности названнаго элемента опредѣляется воззрѣніе на различныя структуры, обнаруживающія при поверхностномъ осмотрѣ нѣкоторое сходство съ тѣмъ, что имѣется у Passeres (вильчатость передняго отдѣла сошника), но совсѣмъ не удовлетворяющія эмбріологическому критерію въ опредѣленіи эгитогнатизма. Отсюда ошибочность или, по меньшей мѣрѣ, за отсутствіемъ онтогенетическихъ указаній — преждевременность всѣхъ вышеупомянутыхъ сопоставленій (Newton, Garrod и др.), какъ и вообще малая пригодность мацерированныхъ препаратовъ взрослыхъ особей и неубѣдительность рисунковъ или описаній таковыхъ, а съ другой стороны — необходимость новыхъ поисковъ для рѣшенія вопроса о діагностичности вновь редактированнаго признака [8] , или, иначе говоря, для уясненія того, насколько данныя онтогенеза одного изъ высшихъ представителей отряда пригодны для характеристики всѣхъ остальныхъ Passeriformes.
Само собою разумѣется, что для сравнительно-анатомической провѣрки нашего опредѣленія эгитогнатизма прежде всего необходимо тщательное изслѣдованіе возможно большаго числа Passeriformes въ отношеніи ихъ небно-челюстного аппарата, и что лишь отсутствіе подходящаго матеріала и трудность полученія такового вынуждаютъ насъ искать рѣшенія вопроса въ изученіи литературныхъ данныхъ, заставляетъ въ частности вернуться къ уже упомянутой въ введеніи работѣ Паркера: «On Aegithognathous Birds» и нѣсколько подробнѣе остановиться на нѣкоторыхъ страницахъ этого объемистаго трактата. При этомъ разсмотрѣніе послѣдняго необходимо предпринять въ двоякомъ направленіи соотвѣтственно двоякой цѣли нашего анализа: съ точки зрѣнія вопроса о приложимости онтогенетическаго критерія опредѣленія эгитогнатизма въ ряду Passeriformes и, далѣе, со стороны вопроса о его діагностичности по отношенію къ другимъ отрядамъ. Не трудно видѣть, что мы здѣсь имѣемъ два самостоятельныхъ вопроса и потому намъ представляется желательнымъ возможно рѣзче разграничить ихъ въ послѣдующемъ изложеніи.
Послѣ этой небольшой вступительной замѣтки обратимся къ Паркерову сочиненію и, заручившись эмбріологическимъ опредѣленіемъ эгитогнатизма, посмотримъ, въ какой мѣрѣ подтверждается оно сравнительно-морфологическими данными.
Прежде всего необходимо здѣсь отмѣтить нѣкоторую условность или затруднительность въ оцѣнкѣ описаній и рисунковъ названной работы. Дѣло въ томъ, что, какъ мы уже имѣли случай указать, ошибочные взгляды Паркера по нѣкоторымъ пунктамъ разбираемой структуры (воображаемые всюду «vomerine cartilages» съ ихъ парными окостенѣніями) сильно затемнили другое, болѣе удачное указаніе того же автора на тѣсныя взаимоотношенія сошника и такъ наз. «inturned alinasal wall». Отмѣтивъ совершенно правильно участіе послѣднихъ въ образованіи передняго развилка сошника, равно какъ и наличность двухъ другихъ окостенѣній, «septo-maxillares» по обѣимъ сторонамъ его, авторъ на совершенно исключительномъ по своему богатству матеріалѣ, обнимавшемъ представителей отъ большинства семействъ Passeriformes, вполнѣ послѣдовательно старается найти для названной особенности различныя ступени дифференцировки. Въ результатѣ — установка нѣкотораго числа «Varieties of the Aegithognathous palate», родъ шкалы, поясняющей извѣстную градацію въ развитіи даннаго признака. Въ этой небольшой таблицѣ, вовсе опускаемой одними авторами и въ неизмѣнномъ видѣ, т.-е. безъ всякихъ комментаріевъ перепечатываемой другими, мы находимъ рядъ короткихъ діагнозовъ, опредѣляющихъ главнѣйшія модификаціи интересующаго насъ типа неба. Такихъ модификацій Паркеръ различаетъ три и называетъ ихъ
- Incomplete,
- Complete и
- Compound-Aegithognathism.
Оставляя покамѣстъ въ сторонѣ послѣдній случай (с), получившій названіе «сложнаго» изъ-за его предполагаемой десмогнатической тенденціи и встрѣчаемый далеко не часто, начнемъ съ модификации b, какъ наиболѣе широко представленной въ ряду эгитогнатовъ, наиболѣе для нихъ типичной и спеціализированной. Подраздѣленіе это, какъ уже сказано, было установлено подъ именемъ «Complete» и, по мнѣнію Паркера, въ свою очередь позволяете различить двѣ слѣдующія разновидности:
| Var. 1. — This occurs in some of the lowest harsh voiced «Coracomorphae». The vomers are developed in large vomerine cartilages, which they often only partially ossify; but there osseous tracts are distinct from those of the often bony alinasal walls and turbinals. A small «septo-maxillary», one each side, generally appears limpetlike, on the inturned angle of the alinasal cartilage, but does not run into it; this is well seen in Pachyrhamphus, Pipra and Tamnophilus. |
| Var. 2. — This occurs in an immense group comprising the higher «Coracomorphae» and also the Swifts (Cypselidae). Here all the vomerine bones are grafted upon the nasal wall, and thus the bird loses its primary «schisognathism». |
Опуская первую половину опредѣленія, несостоятельную благодаря введенію ирраціональнаго критерія, посмотримъ, каково различіе другихъ фактическихъ особенностей, поведшихъ къ названному расщепленію.
Къ числу подобныхъ признаковъ, характеризующихъ «the first variety of complete aegithognathism» куда, помимо перечисленныхъ примѣровъ, принадлежатъ строеніе неба и другихъ Tracheophonae: Dendrocolaptes, Synallaxis, Muscisaxicolla, Homorus... относится раздѣльное, обособленное существованіе тѣхъ ингредіентовъ (заднie отдѣлы носового лабиринта, «septo- maxillaria» и собственно передній сошниковый), которые у высшихъ представителей отряда, сливаясь между собою, вызываютъ своеобразную конфигурацію сошника. Другими словами, у нѣкоторыхъ Passeres означенные элементы пребываютъ какъ бы въ сыромъ, несцементированномъ видѣ — въ состояніи, палингенетически-преходящемъ для большинства другихъ Passeriformes. Таково, по крайней мѣрѣ, впечатлѣніе отъ соотвѣтствующихъ таблицъ и описаній обсуждаемой работы, и, беря во вниманіе данныя онтогенеза, изложенныя во второй главѣ нашей статьи, мы въ правѣ усмотрѣть въ этомъ пожизненномъ сохраненіи эмбріонально-примитивныхъ чертъ Tracheophonae филогенетическую стадію эгитогнатизма, удержанную отъ сравнительно недавняго прошлаго. Однако, принимая Паркеровы термины какъ выраженія извѣстной филетической послѣдовательности въ развитіи даннаго признака, мы тѣмъ рѣшительнѣе высказываемся противъ ихъ систематическаго приложенія въ томъ смыслѣ, какъ это пытался сдѣлать Паркеръ: не говоря уже о томъ, что, по свидѣтельству того же автора, въ группѣ Tracheophonae, на ряду съ преобладаніемъ 1-й разновидности полнаго эгитогнатизма, встрѣчается также «complete aegithognathism of the 2-nd variety» (Pitta), и даже случаи эгитогнатизма «of the highest degree» (Grallaria), изученіе другихъ признаковъ, взятыхъ изъ организаціи высшихъ представителей отряда, съ очевидностью показываетъ намъ, что положеніе, количество и размѣры такъ наз. «septo-maxillare» — передне- боковыхъ окостенѣній сошника — весьма непостоянны даже въ тѣсныхъ предѣлахъ семейства или рода. Какъ мы увидимъ ниже, остающееся за вычетомъ примѣровъ «1-й разновидности» эгитогнатизма огромное количество Passeriformes, морфологически объединенное Паркеромъ на основаніи болѣе совершенной, спеціализированной формы небно-челюстнаго аппарата, на дѣлѣ обнимаетъ собою формы, далеко неоднородныя въ отношеніи небно-лицевой структуры, при чемъ размѣры этого взаимнаго несходства даже превосходятъ тѣ отличія, которыя повели къ установленію указанныхъ двухъ разновидностей. Однако, прежде чѣмъ приступить къ разбору этой 2-й разновидности «of the complete Aegithognathism», остановимся попутно на нѣкоторыхъ другихъ особенностяхъ черепа Passeriformes, впервые выдвинутыхъ Паркеромъ, какъ дополнительный моментъ при различеніи «высшаго» и «низшаго» типа эгитогнатизма.
Мы видѣли, что неодинаковая степень слитія отдѣльныхъ компонентовъ сошника и, въ частности, различное соединеніе «septo-maxillare» съ прилежащими частями хрящевого лабиринта — послужили главнымъ поводомъ къ установленію двухъ категорій полнаго эгитогнатизма, въ томъ видѣ, какъ они формулированы въ Паркеровой таблицѣ, что же до ихъ обоснованія въ текстѣ, то оно, какъ и обычно, сопровождается у Паркера большимъ количествомъ апатомическихъ деталей, совершенно затемняющихъ искомую аргументацію. Однако, въ числѣ такихъ подробностей, упоминаемыхъ попутно въ текстѣ при отдѣльныхъ описаніяхъ и не вошедшихъ въ названную таблицу, мы находимъ нѣкоторыя черты строенія лицевого черепа, отчасти подтверждающія воззрѣнія автора «Оn the Aegithognathous Birds» и потому заслуживающія быть здѣсь приведенными. Сопоставляя относящіяся сюда замѣтки Паркера, разбросанныя на протяженіи болѣе ста страницъ, мы получаемъ слѣдующій рядъ признаковъ лицевого скелета, въ различной мѣрѣ дополняющихъ характеристику обѣихъ разновидностей «полнаго» эгитогнатизма.
Размѣры «Transpalatina» ... [ихъ зачаточность у низшихъ формъ].
Степень выраженности Proc. maxillo-palatini ... [отсутствіе срединнаго перехвата какъ указаніе примитивности].
Конфигурація Praepalatina ... [первоначальная прямизна ихъ, филетически позднѣйшая изогнутость].
Форма Pterygoidea ... [ихъ прогрессивное утонченіе въ ряду Passeres].
Величина и способъ окончанія Postpalatina ... [болѣе значительная величина и скошенность у примитивныхъ формъ, укороченность и заостренность у высшихъ представителей].
Степень схожденія Postpalatina ... [сильное схожденіе характеризуетъ низшихъ членовъ группы].
Развитіе Interpalatina ... [относительно бо́льшее у низшихъ представителей Passeres].
Степень схожденія Ethmopalatina за уровень предыдущихъ выростовъ ... [отсутствіе такового характерно для многихъ примитивныхъ формъ].
Присутствіе Lacrymale ... [свойственно преимущественно высшимъ представителямъ].
Способъ выхожденія нервовъ 1 и 5 пары ... [общность выходныхъ отверстій — состояніе примитивное, раздѣльность — признакъ спеціализаціи].
Совершенно оставляя въ сторонѣ другіе признаки (напр., существованіе Proc. epipterygoideus, наличность Fissura cranio-facialis, присутствіе и положеніе Proc. basipterygoidei, размѣры носовыхъ окостенѣній), вовсе непригодные для классификаціонныхъ цѣлей, необходимо указать, что и для вышеперечисленныхъ морфологическія толкованія не одинаково опредѣленны. Такъ, нѣкоторыя черты, въ дефинитивномъ состояніи регулярно находимыя у формъ, примитивныхъ по совокупности своей организаціи, наблюдаются временно у другихъ представителей отряда, въ цѣломъ болѣе спеціализированныхъ, и въ этомъ смыслѣ позднее образованіе Transpalatina, позднее же пріобрѣтеніе характерной формы небно-челюстныхъ отростковъ, Palatina и Pterigoidea, — замѣщающихъ палингенетическія простыя отношенія тѣхъ же элементовъ на болѣе ранней стадіи зародыша (см. Глава 1, Фактическія основанія классификаціи Гексли и позднѣйшая ихъ критика главу настоящей работы) являются существеннымъ таксономическимъ моментомъ, позволяющимъ въ исторіи развитія одного изъ представителей Corvidae видѣть отчасти «the measure of the rest». Напротивъ, что касается другихъ упомянутыхъ особенностей, то, за отсутствіемъ или неясности «біогенетическихъ» указаній, ихъ филогенетическій возрастъ, а слѣдов. и значеніе для систематики гораздо менѣе очевидны. Поэтому, не отрицая за подобными различіями въ строеніи небно-крыловидно-челюстного комплекса извѣстнаго морфологическаго интереса, какъ подтверждающими мысль о сравнительно недавнемъ происхожденіи нѣкоторыхъ второстепенныхъ чертъ, характерныхъ для высшей формы эгитогнатизма, и слѣдов. О дифференцировкѣ этихъ послѣднихъ внутри самихъ Passeriformes, мы оставляемъ здѣсь открытымъ вопросъ о собственно классификаціонномъ ихъ значеніи и переходимъ къ разсмотрѣнію слѣдующей, второй по счету, разновидности интересующей насъ формы неба.
Нѣсколькими строками выше, говоря о черепѣ Tracheophonae, «the lowest harsh voiced Coracomorphae» Паркера, мы старались показать, что отличительное свойство ихъ небно-челюстнаго аппарата, пожизненно нетѣсное соединеніе элементовъ сошника, характеризуя временное состояніе неба у птенцовъ «the higher Coracomorphae», тѣмъ самымъ побуждаетъ насъ признать за этой формой эгитогнатизма у Tracheophonae значеніе извѣстной филетической ступени въ развитіи неба у Passeriformes, при этомъ совершенно независимо отъ примѣненія ея въ классификаціонномъ направленіи. Посмотримъ, не удается ли и въ отношеніи другихъ, оставшихся примѣровъ «On the Aegithognathous Birds» притти къ аналогичнымъ результатамъ, т.-е. установить для наиболѣе характерной части ихъ черепа, сошника и смежныхъ элементовъ, различій филогенетическаго свойства, или, выражаясь иначе, найти такіе признаки и состоянія, которые помогали бы намъ установить сравнительно-анатомически главнѣйшіе моменты ея морфологической дифференцировки.
Къ числу наиболѣе рельефныхъ признаковъ подобнаго рода относится строеніе сошника Menura, — формы, послужившей, какъ извѣстно, темой безконечныхъ разногласій орнитологовъ со дня открытія и вплоть до появленія «On the Classification of Birds». Авторъ послѣдней въ свое время не преминулъ отмѣтить своеобразное устройства сошника у лирохвоста, напоминающее эгитогнатовъ своей глубокой задней расщепленностью, но отличающееся отсутствіемъ передняго развилка. Различіе это, несмотря на поврежденность экземпляра, бывшаго въ распоряженіи Гексли, не въ состояніи было затемнить значеніе конфигураціи всѣхъ прочихъ лицевыхъ костей и общей формы черепа, не помѣшало «to divide the Coracomorphae into two primary groups — one containing Menura, and the other all the other genera, which have yet been examined» (p. 472). Согласно этому, и въ обсуждаемой работѣ Паркера подъ рубрикой «Оn the Morphology of the Face in the Coracomorphae» одна изъ первыхъ монографій посвящена скелету головы этой интересной австралійской птицы. Авторъ подробно останавливается на ея лицевомъ скелетѣ и, такъ сказать, на фонѣ безусловнаго признанія эгитогнатическаго habitus'а небно-челюстного аппарата, отмѣчаетъ рядъ болѣе простыхъ и примитивныхъ чертъ въ расположенiи и формѣ нѣкоторыхъ отдѣльныхъ элементовъ (напр., общее отверстіе для выхожденія нервовъ I иV пары, малое развитіе ethmopalatina), и на первомъ мѣстѣ — любопытное устройство сошника. Это — массивная, сверху выемчатая пластинка, кзади вытянутая въ «crura vomeris», спереди ланцетовидно-заостренная безъ всякаго слѣда рожковъ, столь характерныхъ для эгитогнатовъ. Что касается другой особенности названнаго типа, болѣе глубокой и общераспространенной, именно тѣснѣйшаго взаимоотношенія носового лабиринта и сошника, то на существованіе такового указываютъ два небольшіе островка хряща — остатки гіалиновыхъ вестибулярныхъ стѣнокъ, изображенные въ связи съ дорсальвыми краями послѣдняго (Табл. B.3, fig. 5. ial). Къ сожалѣнію, всѣ относящіеся сюда рисунки (фиг. 1—5) Паркера изображают черепъ лирохвоста въ мацерированномъ [9] видѣ, съ частями хрящевого лабиринта совершенно удаленными, и потому объ истинной морфологической природѣ этихъ гіалиновыхъ участковъ, представленныхъ на мѣстѣ, соотвѣтствующемъ заднимъ, внутрь загибающимся частямъ вестибулярныхъ стѣнокъ лабиринта, мы, вмѣстѣ съ Паркеромъ, догадываемся лишь на основаніи аналогіи съ высшими Passeriformes. Съ другой стороны, второе предположеніе того же автора о вѣроятномъ присутствіи лабиринтарныхъ костныхъ центровъ по бокамъ отъ вершины сошника, конечно, совершенно произвольно.
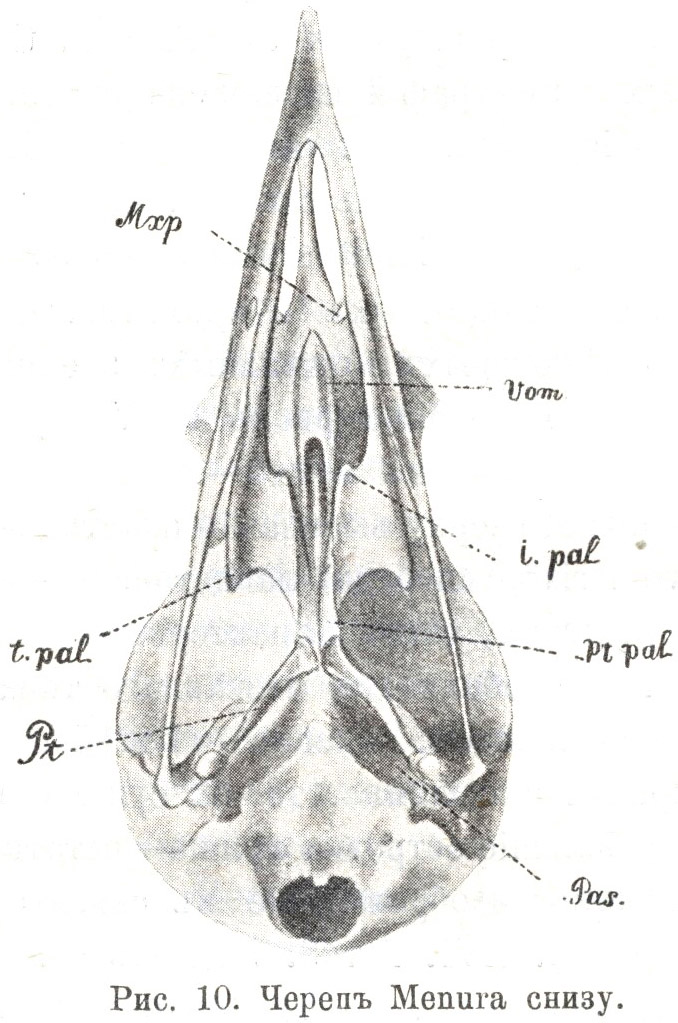
Однако, если ограничиться разсмотрѣніемъ послѣдняго
элемента въ отдѣльности, безъ отношенія къ степени
участія прилежащихъ хрящевыхъ отдѣловъ лабиринта въ
построеніи его передняго конца, то необходимо согласиться, что
Паркеръ совершенно правильно отмѣтилъ его кажущуюся
недоразвитость по сравненію съ тѣмъ, что наблюдается у
высшихъ Passeres, равно какъ и соотвѣтствiе между дефинитивными
очертаніями сошника Menura и зародышевымъ состояніемъ того
же элемента у развивающагося Corvus. Въ самомъ дѣлѣ,
достаточно одного взгляда на прилагаемый здѣсь рисунокъ черепа
Menura (Табл. 3.1) и бѣглаго сравненія съ
помѣщеннымъ на Табл. B.1, fig. 39 нашей работы,
изображеніемъ неба молодого Corvus frugilegus, чтобы увидѣть
поразительное сходство въ строеніи сошника того и другого вида.
Такимъ образомъ, организація Menura, въ отношеніи
интересующаго насъ отдѣла черепа, позволяетъ намъ вернуться еще
далѣе назадъ въ исторіи его филогенеза и возстановить
сравнительно-анатомически одну изъ первоначальныхъ ступеней его, и въ этой
морфологической оцѣнкѣ
признака мы, какъ и въ предыдущіе раза, при обсужденіи черепа
Tracheophonae, — согласны съ мнѣніемъ
англійскаго зоолога. Съ другой стороны, здѣсь въ еще гораздо
большей мѣрѣ обнаруживается непригодность разбираемаго
признака для собственно классификаціонныхъ цѣлей, какъ
критерія оцѣнки высоты организаціи,
діагностичнаго въ предѣлахъ меньшихъ группъ, семейства или
подотряда. Насъ убѣждаютъ въ этомъ дальнѣйшія
наблюденія того же автора, до сихъ поръ оставлявшіяся, правда,
безъ вниманія, именно, указанія того, что въ группѣ,
характеризуемой in toto принадлежностью къ
высшей формѣ эгитогнатизма (т. наз. 2-ой разновидности), спорадически
находили формы, которыя со стороны изслѣдуемаго признака —
отсутствіемъ передне-сошниковаго развилка напоминаютъ
состояніе Menura.
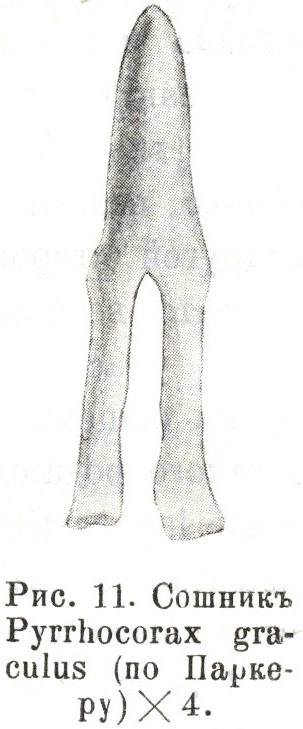
Дѣйствительно, какъ слѣдуетъ изъ
Паркеровыхъ рисунковъ и описаній сошника отъ Pyrrhocorax graculus (22. I. 307. Pl. LV. fig. 10-12),
ближайшіе родичи котораго, являясь эгитогнатами par excellence,
повторно избирались въ качествѣ примѣра наивысшей
спеціализированности названнаго типа, мы здѣсь, у клушицы
(Табл. 3.2), во взросломъ состояніи, находимъ ту же
эмбріональную конфигурацію этой кости, что и у лирохвоста,
формы, — отнесенной большинствомъ анатомовъ далеко къ
основанію генеалогической вѣтви Passeres. Такое
нахожденіе примитивныхъ чертъ въ строеніи черепа у одного изъ
высшихъ представителей отряда, какъ далѣе оказывается, отнюдь не
представляется единичнымъ; оказывается, что въ тѣсныхъ
предѣлахъ одного и того же семейства, вдобавокъ группы, издавна
помѣщаемой во главѣ Passeriformes, случаи недоразвитія
передне-боковыхъ окостенѣній («septo-maxillaria»)
сошника и болѣе обычные примѣры ихъ существованія
встрѣчаются бокъ о бокъ, совершенно спорадично. По крайней
мѣрѣ на стр. 305 обсуждаемой работы, при описаніи неба
Ruticilla phoenicura, мы находимъ
слѣдующее признаніе Паркера: «Neither in the yonng
Redstart, nor in the adult Whitethroat (Sylvia
cinerea) have I been able to detect any lateral ossicles or septo-
maxillaries. I have also searched for them in vain in the Wagtails (Budytes rayi and Motacilla yarrelli); but in the Willowwren
(Phylloscopus trochilus) they are very evident
on each side in the substance of the nasal cartilage. In the Redbreast (Erithacus rubecula) they are very small»...
Согласно этому, и вильчатость передняго отдѣла сошника въ различной
мѣрѣ приближается къ тому, что установлено для Pyrrhocorax,
такъ, напр., у Ruticilla (Pl. LV, fig. 13) съ ея сравнительно простымъ переднимъ
сошниковымъ краемъ, — и допуская даже, что въ случаѣ Menura
мы дѣйствительно имѣемъ филогенетически-первоначальныя
отношенія въ противоположность Pyrrhocorax, Sylvia и Ruticilla съ ихъ
вторичнымъ упрощеніемъ, благодаря задержкѣ и редукціи
означенныхъ лабиринтарно-сошниковыхъ центровъ, мы, тѣмъ не
менѣе, за внѣшней сходностью обоихъ состояній и при
отсутствіи критерія для выясненія ихъ истинной природы
въ каждомъ частномъ случаѣ, можемъ лишь отмѣтить фактъ
непостоянства самаго признака и его неприложимость въ систематикѣ съ
точки зрѣнія дифференціативнаго момента.
Если, такимъ образомъ, присутствіе лабиринтарнаго развилка сошника нельзя назвать чертой, всеобщей для эгитогнатовъ, то, съ другой стороны, изъ наблюденій того же Паркера слѣдуетъ допустить возможность образованія вильчатой вершины сошника, помимо участія его лабиринтарныхъ компонентовъ. Наблюденія эти, въ общемъ довольно сбивчивыя и противорѣчивыя, относятся къ такого рода случаямъ, когда [10] собственно сошниковые элементы («vomerine cartilages!») «аrе themselves large enough to form a substratum for all the outgrowings of the vomer»... — слѣдовательно, безъ всякаго участія носовыхъ хрящей, или [11] когда участіе septo-maxillare въ построеніи развилка сошника сравнительно ничтожно. Оставляя въ сторонѣ вопросъ о достовѣрности этихъ указаній, сомнительныхъ не только вслѣдствіе ошибочныхъ воззрѣній автора на сущность эгитогнатизма, постоянно приводимыхъ описаній элементовъ, несомнѣнное отсутствіе которыхъ нами было констатировано для Corvus frugilegus, — но и въ виду чрезвычайной трудности, опредѣлить макроскопически, безъ помощи разрѣзовъ, двойственный составъ эгитогнатическаго сошника; далѣе, помимо бо́льшей вѣроятности иного толкованія, а именно того, что въ приведенныхъ случаяхъ мы имѣемъ дѣло съ временной задержкой въ появленіи добавочныхъ (лабиринтарныхъ) окостенѣній сошника или, напротивъ, съ преждевременной ассимиляціей ихъ со стороны послѣдняго, — можно сказать, что общее значеніе этихъ и подобныхъ наблюденій лишь подтверждаетъ смыслъ предыдущей категоріи фактовъ. Тѣ и другіе говорятъ, что характерныя для большинства эгитогнатовъ лабиринтарныя окостенѣнія сошника, «Septo-maxillare» Паркера, не представляютъ вовсе морфологически самостоятельныхъ образованій, но, связанныя вторично съ костнымъ веществомъ послѣдняго, при случаѣ могутъ и недоразвиться, что при оцѣнкѣ эгитогнатическаго неба главное вниманіе должно быть обращаемо не столько на участіе вестибулярнаго отдѣла въ построеніи извѣстнаго покровнаго элемента (какъ это предлагалось нами во второй главѣ нашей статьи, на основаніи эмбріологическихъ данныхъ) и не на исчисленіе наблюдаемыхъ при этомъ островковъ окостенѣнія, на положеніе, количество, и размѣры Septo-maxillaria (какъ это дѣлалъ Паркеръ въ своей сравнительно-анатомической классификаціи подотдѣловъ типа), сколько на черту, присущую рѣшительно всѣмъ Passeriformes, когда-либо изслѣдованнымъ въ отношеніи небно-челюстного аппарата, а именно на положеніе задняго отдѣла лабиринта, точнѣе заднихъ, медіально загнутыхъ частей вестибулярныхъ стѣнокъ, прилежащихъ непосредственно къ передне-боковому краю сошника. Иначе говоря: если для огромнаго большинства Passeriformes приложимо то опредѣленіе эгитогнатизма, къ которому насъ привела исторія онтогенеза Corvus frugdegus — и которое сводилось къ выдвиганію сложной, двойственной природы сошника, то констатированное Паркеромъ недоразвитіе лабиринтарныхъ компонентовъ сошника у нѣкоторыхъ формъ (Pyrrhocorax, Ruticilla) и менѣе достовѣрные случаи образованія его передняго развилка безъ участія носовыхъ хрящей (Thamnophilus, Dendrocolaptes) побуждаютъ насъ расширить наше первоначальное опредѣленіе въ томъ смыслѣ, что не въ очертаніи вершины сошника — всегда глубоко расщепленной сзади и дорсально выемчатой пластинки, и не въ способѣ ея образованія, но въ тѣсной вообще топографической связи ея съ частями носового лабиринта заключается неизмѣнно общее свойство эгитогнатическаго типа.
Изученіе главнѣйшаго отдѣла Паркеровой группировки, отдѣла «полнаго эгитогнатизма» съ его двумя подраздѣленіями — var. 1&2, привело насъ такимъ образомъ къ рѣшенію одного изъ двухъ вопросовъ, поставлевныхъ нами выше: къ вопросу о приложимости онтогенетического критерія при опредѣленіи эгитогнатизма въ предѣлахъ Passeriformes. Отвѣтъ получился отрицательный въ формѣ приведенной только что поправки на прежнее эмбріологическое опредѣленіе термина. Въ то же время мы старались показать, что выдѣленіе особыхъ разновидностей «varieties of the Complete Aegithognathism», мало пригодное для собственно классификаціонныхъ цѣлей, для уясненія взаимныхъ отношеній различныхъ группъ, условно можетъ быть удержано, какъ выраженіе двухъ послѣдовательныхъ стадій въ развитіи извѣстнаго морфологического состоянія.
Покончивъ съ первой половиной нашей задачи, переходимъ ко второй — къ вопросу о преемственности выдвигаемаго эгитогнатизма за предѣлами Passeriformes, и для выясненія его мы обратимся къ разсмотрѣнію другого, меньшаго, отдѣла Паркеровой группировки — къ разбору понятія «неполный эгитогнатизмъ».
Установленіе этой модификаціи типа было вызвано повѣрочнымъ изслѣдованіемъ небольшой, но своеобразной группы Hemipodii, въ свое время отнесенной Гексли къ шизогнатамъ, на дѣлѣ же обнаруживающей любопытную близость къ Passeres въ строеніи костнаго и хрящевого неба. Въ указанной таблицѣ Паркера «Incomplete Aegithognathism» опредѣляется слѣдующимъ образомъ:
The vomerine cartilages (cartilages to which the symmetrical vomers are attached) are very large, and incomplitely ossified, and the broad double vomer has a «septo-maxillary» at each angle; but these bones are only strongly tied to the «alinasal» cartilage, and do not graft themselves upon it: their union is with the vomerine cartilage.
Здѣсь, какъ и въ предыдущемъ случаѣ, прежде всего приходится отмѣтить несовмѣстимость даннаго опредѣленія съ тѣмъ, что было установлено во 2-ой главѣ нашей работы. Не говоря уже о выдвиганіи на первый планъ размѣровъ и формъ образованій, совершенно отсутствующихъ у Corvus frugilegus и болѣе чѣмъ спорныхъ въ отношеніи другихъ представителей класса, подчеркнутая курсивомъ парность сошника рѣшительно ничѣмъ не подтверждается, а нѣсколько отличному соединенію Septomaxillare съ прилежащими частями лабиринта едва ли можно придавать особое значеніе, въ виду значительной измѣнчивости этихъ отношеній у типичныхъ эгитогнатовъ, — измѣнчивости, доходящей, какъ мы видѣли, до полной утраты этихъ лабиринтарно-сошниковыхъ окостенѣній. Но трудно видѣть, что при такой формулировкѣ, насквозь проникнутой смѣшеніемъ субъективныхъ и повидимому ложныхъ [12] представленій о «хрящевой» закладкѣ сошниковыхъ элементовъ и описаній нѣкоторыхъ дѣйствительныхъ, хотя и маловажныхъ признаковъ, представляется довольно затруднительнымъ составить себѣ должную картину подлинныхъ соотношеній. Если, за отсутствіемъ надлежащей точности въ опредѣленія термина, мы обратимся къ слѣдующимъ затѣмъ страницамъ (р. 295, 296), посвященнымъ болѣе детальному изложенію предмета, то и здѣсь, оказывается, дѣло обстоитъ не лучше. Повидимому, авторъ старается подчеркнуть малое развитіе «Septo-maxillare», тѣхъ парныхъ небольшихъ окостенѣній, которыя у высшихъ Passeres нормально идутъ на развитіе сошниковаго развилка, путемъ ассимиляціи прилежащаго отдѣла носового лабиринта и какъ бы связываютъ его съ вершиной сошника, тогда какъ у Hemipodius и Тurnіх, лежа по бокамъ отъ Vomer, не захватываютъ вовсе лабиринтарной области. Дѣйствительно, на соотвѣтствующихъ рисункахъ (Табл. B.1) части лабиринта не доходятъ до передне-бокового края сошника, будучи отдѣлены отъ него извѣстнымъ промежуткомъ, — состояніе, отличное отъ того, что мы имѣемъ при типичномъ эгитогнатизмѣ. Съ другой стороны, судя по таблицѣ, очертанія сошника (ширина и характеръ передняго окончанія) и вся конфигурація нижней черепной поверхности дѣйствительно напоминаетъ то, что въ болѣе рѣзкой формѣ наблюдается у Passeres. Въ остальномъ однако, иллюстративная часть этой главы очень мало инструктивна, и если только изображенная на рис. 8 причудливая форма «сошниковыхъ хрящей» не безсознательно-искусственный продуктъ извѣстной препаровки, то одного присутствія послѣднихъ было бы достаточно, чтобы исключить всякую мысль о возможности сближенія Hemipodii съ настоящими эгитогнатами. Повторное изученіе краніологическихъ особенностей этой интересной группы является поэтому въ высокой степени желательнымъ, а до тѣхъ поръ вопросъ объ эгитогнатизмѣ Hemipodii, за неудовлетворительностью описанія его у Паркера, приходится считать открытымъ.
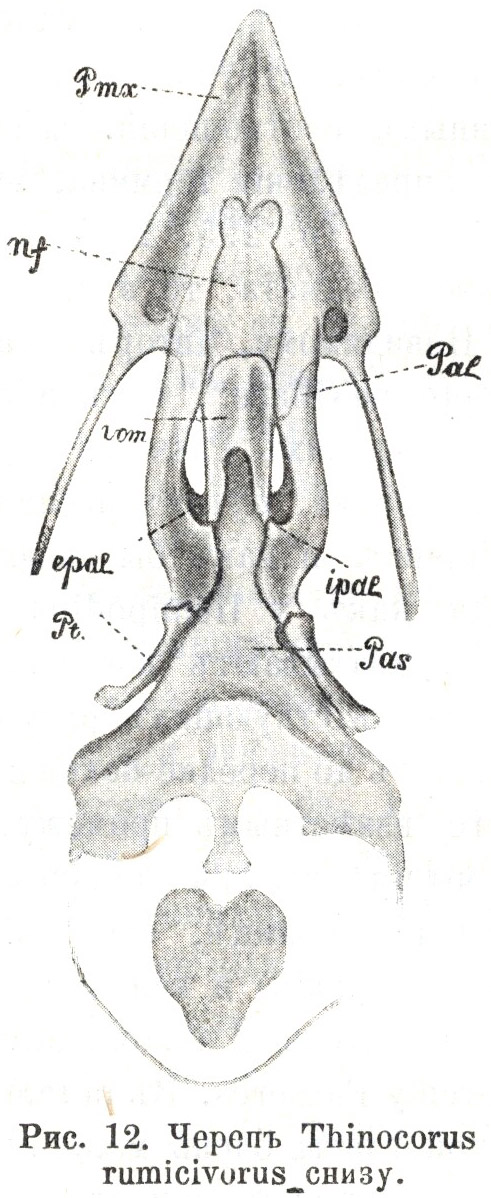
Тѣмъ большее вниманіе заслуживаете небольшой отдѣлъ обсуждаемой работы, которымъ завершается рядъ монографій по эгитогнатамъ. Въ этомъ послѣднемъ, 70-мъ по счету, очеркѣ мы находимъ описаніе неба одного изъ представителей Thinocoridae и вмѣстѣ съ тѣмъ единственное безспорное свидѣтельство существованія типа неба Passeres за предѣлами этой обширной группы, въ строеніи черепа казавшейся дотолѣ менѣе замкнутой, чѣмъ въ отношеніи остальной организаціи. Здѣсь, въ описаніи небно-лицевого аппарата у Thinocorus rumicivorus (22, Part. II, p. 301—306), — описаніи, не затемненномъ ссылкой на загадочные vomerine cartilages (послѣдніе на этотъ разъ не найдены и самимъ авторомъ), — мы впервые (послѣ Passeres) наталкиваемся на то характерное свойство, которое мы признали наиболѣе существеннымъ отличіемъ эгитогнатизма: тѣсную связь между вершиной сошника и частями носового лабиринта. Въ самомъ дѣлѣ, обращаясь къ соотвѣтствующимъ рисункамъ на таблицѣ Паркера (Табл. 3.3, Табл. 3.4), мы видимъ широкій и пластинчатый сошникъ, немного выемчатый сверху, снизу вытянутый въ продольный медіальный гребень, по линіи срастанія правой и лѣвой сошниковыхъ половинъ, не связанныхъ первоначально; послѣднія на протяженіи задней своей трети остаются раздѣленными, образуя на сформировавшейся кости глубоко вдающійся развилокъ. Что касается передняго края сошника, то у различныхъ особей того же вида онъ представляется неодинаковымъ, вѣроятно въ зависимости отъ возраста различныхъ экземпляровъ, а именно, въ различной мѣрѣ скошеннымъ и закругленнымъ, что, впрочемъ, отнюдь не затемняетъ общаго характера элемента, лишь съ внѣшней стороны сравнимаго съ той уклоняющейся формой сошника, которая спорадично была найдена (Garrod) у нѣкоторыхъ представителей этого отряда, и глубоко отличнаго отъ того, что представляетъ большинство другихъ шизогнатическихъ Limicolae. Сошникъ Thinocorus существенно ничѣмъ не отличается отъ сошника отдѣльныхъ Passeres, напоминая случаи, когда, за недоразвитіемъ окостенѣній лабиринтарныхъ боковыхъ частей, сошникъ является простой костяной пластинкой, безъ всякаго слѣда передняго развилка (Menura, Pyrrhocorax, Ruticilla), и, такъ же, какь у настоящихъ Passeres, задніе отдѣлы носового лабиринта, въ видѣ парныхъ хрящевыхъ полосокъ, прилегаютъ непосредственно къ вершинѣ сошника, подстилая его передне-боковыя части.
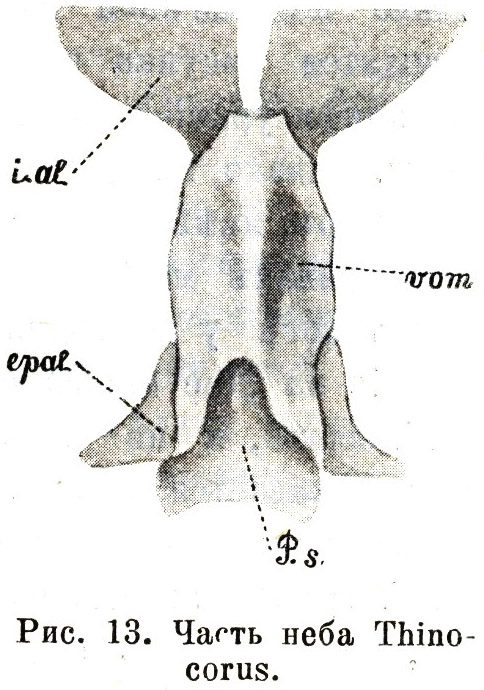
Къ сожалѣнію, за неимѣніемъ
соотвѣтствующаго матеріала, мы лишены возможности
провѣрить нѣкоторыя стороны Паркерова описанія
[13]
; но, если даже допустить извѣстныя отличія въ
конфигураціи частей носового лабиринта, самый фактъ
тѣснѣйшаго взаимоотношенія двухъ элементовъ, столь
различныхъ со точки зрѣнія морфологической природы и
первоначальной ихъ закладки, придаетъ строенію черепа Thinocorus
совершенно исключительный интересъ, и, принимая во вниманіе то, что
за предѣлами Passeriformes мы въ первый разъ находимъ признакъ,
единственно объединяющій всѣхъ членовъ этого обширнаго
подраздѣленія, что, далѣе, его присутствіе у
Thinocorus сопровождается наличностью другихъ особенностей черепа
(очертаніе надклювья, форма Palatina и Pterygoidea, отсутствіе
proc. basipterygoidei, отсутствіе боковыхъ затылочныхъ фонтанелей и
др.), характерныхъ, хотя и въ меньшей степени, для эгитогнатовъ, и большей
частью негативныхъ въ отношеніи собственно Limicolae, что, наконецъ,
за вычетомъ сомнительнаго показанія Паркера объ эгитогнатизмѣ
Turnix, намъ не извѣстны случаи нахожденія названнаго признака
внѣ характеризуемаго имъ отряда Passeres, — нельзя не удивиться,
что любопытное строеніе неба у Thinocorus такъ мало обратило на себя
вниманіе послѣдующихъ авторовъ и въ подробностяхъ осталось
не изученнымъ. Такъ, напр., въ изслѣдованіи Garrod'а: Notes on
the Anatomy and Systematic position of the genera Thinocorus and Attagis (83),
единственной работѣ, спеціально посвященной изученію
этихъ уклоняющихся Limicolae, приводятся рисунки черепа Attagis,
напоминающаго предыдущій видъ конфигураціей надклювья,
небно-лицевыхъ костей и шириною Vomer. Однако, въ противоположность
объекту Паркерова описанія, экземпляръ, имѣвшійся въ
распоряженіи Garrod'a, былъ, очевидно, препарированъ
мацераціоннымъ способомъ, соотвѣтственно чему и въ
текстѣ у послѣдняго мы находимъ (р. 416) только общее
указаніе на «the almost exact identity» въ строеніи
неба у Attagis и Thinocorus, и ни слова объ интересующихъ насъ деталяхъ. Что
же касается позднѣйшей литературы — мы разумѣемъ рядъ
общеизвѣстныхъ орнитологическихъ компендіевъ, — то въ
отношеніи данной группы авторы ихъ всецѣло опираются на
показанія Паркера, довольствуясь формальной ссылкой на
«incomplete Aegithognathism of Attagis and Thinocorus» и
воздерживаясь отъ какихъ бы то ни было поправокъ или поясненій. Но
даже здѣсь, при усвоеніи Паркеровой терминологіи,
принявшіе ее руководились исключительно одной чертой, именно,
формой сошника — чертой, сравнительно второстепенной по
сравненію съ другой особенностью черепа Thinocoridae —
положеніемъ и устройствомъ хрящевого лабиринта, игнорируемой
совершенно. Такъ, Beddard, давшій наиболѣе оригинальную,
пополненную собственными наблюденіями, сводку фактовъ,
относящихся къ вопросу о таксономическомъ значеніи небно-лицевого
аппарата птицъ, въ систематической части своей книги, называя черепъ у
Thinocoridae «aegithognathous rather then schizognathous» (p. 349),
ссылается на помѣщенныя немного выше (р. 338) пару строкъ: «In
Thinocorus and Attagis the vomer is short and broad, and almost passerine in
front», а въ анатомическомъ отдѣлѣ (p. 150), указывая
бѣгло на описанное Паркеромъ «the union of the vomer with the
ossified alinasals» у Turnicomorphae, — описаніе
чрезвычайно сбивчивое и мало доказательное, не упоминаетъ вовсе объ
аналогичномъ и гораздо болѣе безспорномъ наблюденіи надъ
Thinocorus. Равнымъ образомъ и остальные авторы — Gadow (2, I, 991),
Newton (12, 878) и даже Fürbringer (5, II, 1032) неизмѣнно повторяютъ
формулировку Паркера, совершенно не касаются вопроса о лабиринтарныхъ
компонентахъ небно-челюстного аппарата у Thinocorus и ничѣмъ
рѣшительно не обнаруживаютъ признанія или оцѣнки
столь замѣчательной находки автора «Оn the Aegithognathous
Birds».
─────── |
Кончая разсмотрѣніе Паркеровой работы «On the Aegithognathous birds», нелишнимъ будетъ резюмировать ходъ нашего аналіза и подвести его главнѣйшіе итоги. Исходя изъ строгаго разграниченія двухъ вопросовъ: о безусловной свойственности признака извѣстной группѣ (или, что то же, о его діагностичности) и, во-вторыхъ, о находимости того же признака внѣ этой группы (или, что то же, о его преемственности за предѣлами послѣдней); пытаясь далѣе провести различіе между таксономическимъ достоинствомъ того или иного признака и его сравнительно-анатомическимъ значеніемъ, мы начали съ обсужденія перваго вопроса и на разборѣ «полной» формы эгитогнатизма признали за ея двумя подраздѣленіями (Var. 1 & 2 of the complete Aegithognathous palate) лишь выраженіе двухъ послѣдующихъ филетическихъ ступеней извѣстнаго морфологическаго состоянія.
Мы указали въ то же время на причины, не позволяющія намъ истолковать таксономически структурныя различія этихъ обѣихъ разновидностей или, что то же, использовать въ систематическомъ отношеніи бо́льшую часть работы Паркера, какъ со стороны текста — въ виду неясности руководящихъ взглядовъ автора, такъ и со стороны таблицъ — за неизбѣжной схематизаціей рисунковъ, въ значительной мѣрѣ являющихся отраженіемъ субъективныхъ толкованій автора, и потому мало пригодныхъ для отыскиванія новыхъ, порой чрезвычайно тонкихъ познавательныхъ моментовъ: по крайней мѣрѣ самое тщательное изученіе «Оn the Aegithognathous birds» заставило насъ отказаться отъ задачи, являющейся конечной цѣлью всякаго подобнаго изслѣдованія — переведенія данныхъ морфологическаго описанія на языкъ генеалогическихъ построеній. Съ другой стороны, наша попытка сличеніемъ литературныхъ указаній — въ данномъ случаѣ матеріаловъ, собранныхъ трудами Паркера — сравнительно- анатомически провѣрить и обосновать достоинство критерія, указаннаго данными онтогенеза одного изъ высшихъ представителей Corvidae, показала, что въ ряду Passeriformes измѣненіе конфигураціи передней части сошника, въ виду измѣнчивыхъ размѣровъ и непостояннаго присутствія его передне-боковыхъ окостенѣній, настолько велики, что требуютъ внесенія второго, дополнительнаго корректива въ опредѣленіе термина, измѣненное уже сравнительно съ его первоначальнымъ смысломъ. Въ результатѣ — новая формулировка, при которой главный отличительный моментъ въ строеніи эгитогнатическаго типа черепа переносится на, свойство, общее всѣмъ рѣшителъно Passeriformes и вмѣстѣ съ тѣмъ діагностичное въ отношеніи тѣхъ птицъ, которыя не принадлежали къ послѣднимъ, раздѣляютъ одинъ изъ ихъ второстепенныхъ признаковъ (переднюю вильчатость Vomer), на особенность, лежащую въ основѣ своеобразнаго и сложнаго устройства небно-лицевого аппарата Passeres, а именно, на тѣсную топографическую связь между вершиной сошника и примыкающимъ дорсалъно заднимъ продолженіемъ вестибулярныхъ стѣнокъ лабиринта.
Перейдя къ вопросу о преемственности вновь опредѣляемаго признака «эгитогнатовъ», мы видѣли затѣмъ, что изъ семидесяти съ лишнимъ формъ, монографически изученныхъ въ «Оn the Aegithognathous birds», въ данномъ случаѣ представляютъ интересъ два очерка — первый и послѣдній: описаніе черепа у Hemipodii во вступительной главѣ и заключительная монографія «The Skull of Thinocorus rumicivorus». Оба случая являются примѣрами «неполной» формы эгитогнатизма, и здѣсь, при крайне сбивчивомъ ея опредѣленіи у Паркера, остается въ полной силѣ то, что было сказано по поводу предыдущего отдѣла Паркеровой группировки — о классификаціонной недостаточности признака, чрезвычайно цѣннаго для реставраціи одной изъ примитивно-исходныхъ стадій эгитогнатизма. Это — о значеніи термина «incomplete», какъ такового. Что же до главнаго, интересующаго насъ вопроса — о возможномъ нахожденіи эгитогнатизма за предѣлами Passeriformes, то въ этомъ отношеніи, достоинство двухъ приведенныхъ очерковъ не одинаково. Описаніе черепа Turnicomorphae, наводящее на нѣкоторыя сопоставленія съ Passeres, въ общемь настолько неудовлетворительно, что не допускаетъ надлежащаго использованія. Напротивъ, констатированіе у Thinocorus тѣхъ самыхъ небныхъ отношеній, присутствіе которыхъ мы признали наиболѣе существенной особенностью эгитогнатизма, въ томъ видоизмѣненномъ опредѣленіи этого понятія, къ которому насъ вынудили изученіе онтогенеза неба Passeres и данныя сравнительно-анатомическихъ изслѣдованій Паркера, т.-е. установленіе эгитогнатическаго состоянія черепа у формы, по совокупности своей организаціи отнесенной далеко отъ Passeres, къ шизогнатическимъ Limicolae — нарушаетъ, и притомъ самымъ нагляднымъ образомъ, естественность состава эгитогнатическаго типа.
Малая изученность строенія неба Hemipodii и односторонность въ пониманіи эгитогнатизма у Thinocoridae не помѣшали большинству анатомовъ и систематиковъ воспользоваться этими двумя примѣрами какъ аргументомъ, подрывающимъ діагностичность классификаціоннаго принципа Гексли въ примѣненіи къ Passeres, и можно вообще сказать, что въ этомъ негативномъ толкованіи смысла Паркеровыхъ наблюденій сходятся воззрѣнія всѣхъ авторовъ, такъ или иначе касавшихся вопроса объ естественности эгитогнатическаго типа, этого послѣдняго отдѣла разбираемой системы, долѣе другихъ противостоявшаго научной критикѣ. Исходя изъ болѣе широкой постановки самаго вопроса и болѣе критической оцѣнки матеріала, мы съ своей стороны должны были признать, что отнесеніе Thinocorus къ эгитогнатамъ, въ противоположность аналогичному сближенію послѣднихъ съ Hemipodii, дѣйствительно имѣетъ за собой глубокое фактическое основаніе, гораздо бо́льшее, чѣмъ можно было думать на основаніи поверхностныхъ литературныхъ данныхъ, и въ этомъ смыслѣ заключительная статья въ «Оn the Aegithognathous birds» Паркера, содержащая впервые описаніе черепа отъ своеобразныхъ, уклоняющихся формъ неотропическихъ Limicolae, только закончила собою длинный рядъ изслѣдованій того же автора, приведшихъ, на ряду съ работами другихъ анатомовъ, ко все большему умаленію достоинства критерія Гексли и, наконецъ, къ признанію искусственности разбираемой системы. Не говоря уже о томъ чрезвычайно сдержанномъ пріемѣ, который она встрѣтила при своемъ первомъ появленіи со стороны тогдашняго издателя «Ibis» [14] и позднѣйшаго автора «А Dictionary of Birds», этого наиболѣе убѣжденнаго противника провозглашенныхъ ею принциповъ, во всей послѣдующей литературѣ общеотрицательное отношеніе къ классификаціи Гексли стало возрастать въ той мѣрѣ, какъ постепенно сглаживались границы прежнихъ «типовъ», и современные зоологи, не отрицая совершенно таксономическаго значенія элементовъ, выдвинутыхъ Гексли, но отводя имъ нѣкоторое мѣсто въ ряду другихъ второстепенныхъ признаковъ, высказываются противъ сохраненія «типовъ» въ томъ видѣ, какъ это предполагалось авторомъ «On the Classification». Такъ, напр., въ неоднократно приводимой книгѣ Beddard'a [15] за краткимъ изложеніемъ особенностей черепа, общихъ всему классу, слѣдуетъ подробное перечисленіе тѣхъ, которыя въ различныхъ группахъ представляютъ наибольшія модификаціи, и здѣсь на первомъ мѣстѣ — измѣненія небно-челюстного аппарата. Коснувшись далѣе принциповъ Гексліевой группировки, авторъ, говоря о типахъ, отмѣчаетъ невозможность ихъ строгаго разграниченія: «Neither are any of the subdivisions, except that of dromaeognathaе, really satisfactory from the classificatory point of view. Their inefficiency, however, is rendered harmless by the fact that they are in reality not such hard and fast distinctious as might bo gathered from the foregoing abstract and from textbooks in general» (p. 140). Подобное же мнѣніе высказываетъ Gadow въ анатомическомъ отдѣлѣ «Bronn's Klassen und Ordnungen». Въ заключительномъ пространномъ очеркѣ исторіи развитія онъ пополняетъ данныя главы остеологіи, написанной Selenka, и, присоединяясь къ болѣе ранней критикѣ [16] этого своего предшественника по составленію названнаго компендія, замѣчаетъ (р. 991): «Hiermit fällt der Werth dieser Formationen als Princip für die Einteilung der Vögel in drei oder vier Gruppen». Наконецъ, Fürbringer, отдавая должное геніальной проницательности великаго зоолога и, такъ сказать, идейной сторонѣ его работы, тѣмъ не менѣе, отказывается признать въ провозглашенныхъ имъ принципахъ таксономическій моментъ первостепеннаго значенія и согласно этому, давая сжатый очеркъ названной системы и короткій перечень ея позднѣйшихъ коррективовъ, начинаетъ словами: «Ohne Zweifel ist das Huxley'sche System ein künstliches und zahlreiche später über die Structur des Gaumens gemachte Beobachtungen haben sich ihm nicht günstig erwiesen» (p. 1031).
Итакъ, ближайшее знакомство съ данными литературы показало намъ, что признакъ, характеризующій строеніе небно-челюстного аппарата Passeres и притомъ единственно объединяющій всѣхъ членовъ этой обширной группы, не есть діагностичный, что характеризуемый имъ типъ эгитогнатовъ, подобно предыдущимъ типамъ не удержался въ систематикѣ и, будучи кассированъ, только ускорилъ, довершилъ паденіе классификаціи Гексли. Является вопросъ: каково же отношеніе этого четвертаго и послѣдняго отдѣла названной системы къ тремъ первымъ категоріямъ — типамъ десмо-, шизо- и дромэогнатическому, которые, какъ мы видѣли (см. Глава 1, Фактическія основанія классификаціи Гексли и позднѣйшая ихъ критика главу), еще раньше удалось связать между собою. Представляется задача: исходя изъ факта анатомической взаимной связи всѣхъ четырехъ типовъ, указать точнѣе ихъ взаимоотношенія и, въ частности, для эгитогнатизма опредѣлить возможно ближе родъ морфологической преемственности. Начнемъ съ послѣдняго, тѣмъ болѣе что обсужденію взаимныхъ отношеній прочихъ типовъ была посвящена значительная часть введенія, и обратимся напередъ къ литературѣ. Здѣсь намъ приходится отмѣтить значительное разногласіе авторовъ. Въ то время какъ одни (Gadow, Newton), стоя на точкѣ зрѣнія Гексліева опредѣленія эгитогнатизма, выдвигаютъ на первый планъ конфигурацію вершины сошника и, опираясь на наблюденія Garrod'a о расщепленности ея у Numenius, Recurvirostra, Haematopus и указанія Паркера о передней вильчатости того же элемента у Megalaema и недоразвитія ея у Turnix и Thinocorus, высказываются за взаимное сближеніе шизо- и эгитогнатовъ, другіе присоединяются къ воззрѣніямъ Pycraft'a, который, по установленіи дѣленія всѣхъ птицъ на Palaeo- и Neognathae, на основаніи другихъ соображеній — различія въ соединеніи Vomer съ небно-крыловидными костями — выводитъ эгитогнатизмъ изъ дромэогнатическаго состоянія.
Несомнѣнно, что подобное различіе во взглядахъ явилось результатомъ неустановившихся воззрѣній на дѣйствительную природу эгитогнатизма, обусловленныхъ чрезмѣрной узостью и догматичностью его первоначальнаго опредѣленія, и что при болѣе широкомъ пониманіи термина, мѣсто прежнихъ сбивчивыхъ, одностороннихъ толкованій занимаетъ ясное, фактически единственно возможное опредѣленіе названнаго типа, помогающее намъ въ то же время намѣтить вѣроятный путь его морфологической дифференцировки.
Въ самомъ дѣлѣ, понятый въ первоначальномъ смыслѣ, «эгитогнатизмъ» пришлось бы неминуемо распространить на нѣкоторыхъ Limicolae, каковы Numenius, Recurvirostra, Chionis и Haemalopus, а положеніе послѣднихъ въ ряду типичныхъ, въ отношеніи структуры неба, представителей отряда естественно приводитъ къ мысли о шизогнатическомъ происхожденіи названнаго типа; съ другой стороны, признаніе лабиринтарной природы сошниковаго развилка какъ бы подтверждаетъ правильность такого взгляда, ибо устанавливаетъ связь между зачаточнымъ эгитогнатизмомъ низшихъ Passeres (Menura) и таковымъ же состояніемъ Thinocorus, уклоняющейся формы тѣхъ же шизогнатическихъ Limicolae. Однако, разсуждая такимъ образомъ, мы въ сущности едва ли приближаемся къ рѣшенію проблемы.
При разсмотрѣніи вопроса о происхожденіи какого-либо признака, мы за точку отправленія беремъ его наиболѣе примитивное состояніе, абстрагируя отъ тѣхъ, порой весьма распространснныхъ и характерныхъ, моментовъ, которые являются продуктами новѣйшаго образованія. Если поэтому въ разсматриваемомъ конкретномъ случаѣ мы видимъ, что извѣстный признакъ (присутствіе лабиринтарнаго развилка у вершины сошника), находимый у огромнаго большинства эгитогнатовъ, являясь негативнымъ въ отношеніи всѣхъ не-Passeriformes, тѣмъ самымъ подтверждаетъ принадлежность первыхъ къ одному морфологическому типу, но, съ другой стороны, отсутствуетъ у нѣкоторыхъ формъ и недоразвитъ въ различной мѣрѣ у другихъ, и если за его вторичную, новѣйшую природу говоритъ исторія развитія, онтогенетически поздняя закладка, — то очевидно, что какъ признакъ высшей спеціализаціи, онъ долженъ быть отброшенъ при обсужденіи вопроса о происхожденіи эгитогнатизма, и что оставшіяся за его вычетомъ существенныя черты послѣдняго и представляютъ то, что въ неизмѣненномъ, чистомъ видѣ мы находимъ за предѣлами Passeriformes, у своеобразныхъ представителей неотропическихъ Limicolae. Отсюда ясно, что попыткой уяснить бдижайшій генезисъ строенія неба у Thinocoridae всего лучше подойти къ рѣшенію вопроса о морфологической преемственности эгитогнатическаго типа.
Возникновеніе эгитогнатизма въ томъ видѣ, какъ онъ существуетъ у Thinocorus, можно вообразить себѣ двояко: либо непосредственно выводя его изъ дромэогнатическаго неба, т.-е. минуя стадно шизогнатизма, либо изъ послѣдняго, какъ дальнѣйшую его модификацію. Въ пользу перваго воззрѣнія, на первый взглядъ, довольно трудно привести достаточное основаніе. Такъ, выдвигаемая нѣкоторыми (Паркеръ, Фюрбрингерт) парная закладка сошника, предположительно [17] указанная для Thinocorus и совсѣмъ не наблюдавшаяся у Corvus frugilegus, вообще не можетъ почитаться существеннымъ отличіемъ разбираемаго типа, ибо, даже признавая вторичной непарность этого образованія у грача, въ виду несомнѣнной парности того же элемента у первоначальныхъ предковъ Sauropsida и сохраненія таковой у большинства рептилій и нѣкоторыхъ Dinornis, едва ли можно придавать особое значеніе упомянутому признаку, при чрезвычайной трудности разграниченія закладки морфологически-парнаго элемента отъ не имѣющихъ значенія такового сверхсмѣтнаго оссификаціоннаго центра, вызваннаго чрезмѣрнымъ разрастаніемъ извѣстнаго костяного участка. Остаются: величина, точнѣе ширина, Vomer и отношенія небно-крыловидныхъ элементовъ къ сошнику и основанію черепа, т.-е. существованіе на извѣстной стадіи развитія эгитогнатовъ отношеній, пожизненно сохраняющихся у Ratitae и Tinamus («Palaeognathae» Pycraft'а). Однако эта смѣна двухъ различныхъ состояній, въ своихъ существенныхъ чертахъ, въ формѣ замѣны крыловидно-сошниковой связи соединеніемъ Vomer съ Palatina отнюдь не пріурочена къ одному какому-либо типу, но, характеризуя зародышевыя отношенія трехъ различныхъ типовъ черепа: шизо-, десмо- и эгитогнатовъ свидѣтельствуетъ только объ единствѣ ихъ происхожденія, рисуетъ намъ картину примитивнаго, исходнаго строенія отдаленныхъ предковъ «Neognathae» и ничего не говоритъ о вѣроятныхъ способахъ или путяхъ дифференцировки внутри этихъ послѣднихъ, т.-е. не предрѣшаетъ указанной альтернативы. Что же до размѣровъ, вѣрнѣе, положенія Vomer у Thinocorus, то, разсматриваемая въ отдѣльности, эта особенность нисколько не обязываетъ насъ къ принятію непосредственной дромэогнатической преемственности. Разрастаніе сошника въ горизонтальномъ направленіи спорадично наблюдается у самыхъ разнообразныхъ группъ, характеризуемыхъ in toto вертикальной постановкой сошниковой пластинки и несомнѣнной принадлежностью къ шизогнатическому типу: таковы описанные Garrod’омъ и неоднократно приведенные примѣры изъ Limicolae (Recurvirostra, Hаematopus и Numenius), таковъ изображенный Гексли горизонтально расширенный сошникъ Tetrao urogallus и нѣкоторые другіе случаи. Само по себѣ латеральное разрастаніе тѣла сошника, какъ частный случай измѣненія формы элемента вообще весьма непостояннаго, — процессъ настолько обыденный и несложный, что не требуетъ необходимаго признанія монофилетичности происхожденія и въ этомъ смыслѣ плоскостное расширеніе сошника Thinocorus и нѣкоторыхъ другихъ Limicolae могло явиться филетически новѣйшимъ признакомъ, самостоятельно пріобрѣтеннымъ.
Итакъ, разсмотрѣнныя до сихъ поръ черты строенія, за малой характерностью и спорадичностью распространенія, не позволяютъ категорически отвѣтить на вопросъ о прошломъ эгитогнатизма, и теперь намъ остается разобрать послѣднее и наиболѣе существенное свойство названнаго типа — соединеніе покровной кости, сошника, съ частями органа обонянія. Къ сожалѣнію, обширные остеологическіе матеріалы, собранные послѣдними десятилѣтіями, въ формѣ все продолжавшагося изученія остающихся по мацерацiи костныхъ элементовъ черепа, свидѣтельствуютъ о полной позабытости обширнаго сочиненія Паркера, впервые, хотя, быть можетъ, недостаточно подробно, уяснившаго значеніе хрящевыхъ ингредіентовъ эгитогнатическаго неба. Въ виду такого игнорированія лабиринтарнаго отдѣла черепа у птицъ, мы затрудняемся, по даннымъ орнитологической литературы прослѣдить внутри того или другого типа зачатки любопытнаго соединенія сошника и лабиринта, не можемъ даже составить себѣ приблизительной картины этихъ отношеній въ интересующихъ насъ отрядахъ. Намъ остается методъ чисто-эмбріологическій: руководясь извѣстною послѣдовательностью, въ которой формируются при онтогенезѣ различные отдѣлы эгитогнатическаго неба, опредѣлить ихъ относительно-различный возрастъ, возстановить, въ общихъ чертахъ, порядокъ филетическаго появленія и, сопоставляя это съ данными организаціи прочихъ типовъ, приблизиться къ рѣшенію вопроса. Сознавая нѣкоторую условность этого «біогенетическаго» метода и связанную съ ней проблематичность выводовъ, мы ограничимся указаніемъ главнѣйшихъ фактовъ.
Изъ хода развитія черепа у Corvus frugilegus, изученнаго нами во 2-й главѣ этой работы, далѣе, изъ многочисленныхъ, хотя и бѣглыхъ указаній Паркера относительно строенія неба прочихъ Passeres и, наконецъ, изъ данныхъ организаціи Thinocorus, мы заключаемъ, что у всѣхъ эгитогнатовъ взаимоотношенія сошника и прилежащаго отдѣла носового лабиринта слагаются чрезвычайно рано, задолго до обособленія Hemipteryyoideum и обусловливаемой имъ утраты зародышеваго палеогнатизма; что, въ противоположность позднему образованію сошниковаго развилка насчетъ лабиринтарныхъ боковыхъ окостенѣній, этому сравнительно новѣйшему пріобрѣтенію Passeriformes, основное свойство небно- челюстного аппарата Passeres: соединеніе упомянутой покровной кости съ частями органа обонянія мы должны разсматриватъ какъ признакъ чрезвычайно древній и филетически отнести его далеко назадъ къ гипотетической организаціи отдаленныхъ предковъ нынѣшнихъ дромэогнатовъ.
Таковы данныя онтогенеза и его біогенетическое толкованіе. Подтверждается ли оно устройствомъ и расположеніемъ обонятельнаго органа у современныхъ Palaeognathae? При настоящемъ состояніи нашихъ свѣдѣній мы не можемъ въ точности удостовѣрить это, вслѣдствіе упомянутой уже односторонности новѣйшихъ остеологическихъ изслѣдованій. Правда, въ одной изъ своихъ наиболѣе раннихъ монографій (14) Паркеръ на обширномъ матеріалѣ подробно ознакомилъ насъ съ строеніемъ хрящевого лабиринта у Ratitae; однако, принимая во вниманіе совсѣмъ иную цѣль этой работы и неизбѣжно связанную съ нею субъективность описаній и рисунковъ, мы затрудняемся использовать имѣющіяся въ ней отрывочныя указанія положительнаго свойства. Десятилѣтія, протекшія со дня опубликованія «On the Structure and Development of the Skull in the Ostrich Tribe», ничего не прибавили къ первоначальнымъ показаніямъ Паркера, а малая доступность изучаемыхъ объектовъ затрудняютъ надлежащую провѣрку. Но даже при наличности достаточнаго матеріала, разумѣется, необходимо совершенно отрѣшиться отъ столь явно уклоняющихся и вторично измѣненныхъ формъ, какъ Struthio и обратиться къ болѣе типичнымъ представителямъ Palaeognathae; только въ низшихъ, примитивныхъ отношеніяхъ черепа Dromaeus или Casuarius можно надѣяться раскрыть зачатки высоко-спеціализированнаго строенія эгитогнатическаго неба, найти какія-нибудь указанія на происхожденіе обоихъ типовъ отъ одного общаго корня.
На предыдущихъ страницахъ мы старались выяснить дѣйствительное положеніе эгитогнатизма въ ряду прочихъ типовъ, старались вывести его изъ болѣе простыхъ, первоначальныхъ отношеній. Признавая недостаточнымъ обоснованія нѣкоторыхъ существующихъ воззрѣній, мы высказались въ пользу вѣроятнаго дромэогнатическаго происхожденія небно-челюстного аппарата Passeres, т.-е. пришли къ воззрѣніямъ, аналогичнымъ взгляду Pycraft'a: «The Schizognathous and Aegithognathous palates are both specialized forms derived by modification of the Dromaeognathous type» [18] . Однако, въ противоположность этому нѣсколько бѣгло высказанному взгляду, мы, исходя изъ болѣе внимательной оцѣнки имѣвшихся литературныхъ данныхъ, при болѣе опредѣленной постановкѣ самаго вопроса и другой аргументаціи, попытались наметить нѣкоторыя ступени въ эволюціи указаннаго типа, и къ числу такихъ «морфологическихъ этаповъ» мы отнесли строеніе неба Thinocorus, — состояніе, давно оставленное большинствомъ эгитогнатовъ, лишь немногими удержанное по настоящее время. Иной вопросъ, обязываетъ ли насъ извѣстное морфологическое толкованіе черепа Thinocorus къ опредѣленнымъ классификаціоннымъ выводамъ, т.-е. должны ли мы разсматривать Thinocoridae какъ видоизмѣненныхъ предковъ Passeres или вообще въ какихъ-либо генетическихъ отношеніяхъ къ послѣднимъ.
Здѣсь, какъ и вообще въ тѣхъ случаяхъ, когда по сходству анатомическаго строенія приходится судить о генетическомъ взаимоотношеніи самихъ формъ, необходимо различать двѣ стороны вопроса:
не имѣемъ ли мы дѣло съ кажущейся идентичностью строенія, съ явленіемъ изоморфизма и аналогіи,
возможны ли на основаніи указаннаго признака какіе бы то ни было классификаціонныя построенія.
Обратимся сначала къ первому вопросу, какъ болѣе частному. Начнемъ съ того, что въ разбираемомъ примѣрѣ сходство признаковъ, а именно, строеніе извѣстнаго отдѣла черепа Passeriformes и Thinocorus, дѣйствительно чрезвычайно полное, и далѣе касается оно такихъ особенностей, которыхъ не только біологическое, но даже функціональное значеніе весьма проблематичны, — обстоятельство чрезвычайно важное, принимая во вниманіе систематическую непригодность адаптивныхъ и физіологически важныхъ органовъ. Правда, современные зоологи по большей части лишь условно говорятъ о признакахъ «морфологическихъ», охотно апеллируя къ преходящему незнанію ихъ истиннаго значенія, но при этомъ нерѣдко ссылаются на «приспособленіе» тамъ, гдѣ зависимость между строеніемъ и средой, въ широкомъ смыслѣ этого слова, рѣшительно неуловима [19] и, какъ въ случаѣ структуры неба, a priori мало вѣроятна. Поэтому, при современномъ состояніи нашихъ свѣдѣній, намъ остается лишь признать «морфологическій», неадаптивный характеръ изучаемой структуры, какъ выраженія неизвѣстныхъ физіологическихъ процессовъ, неизслѣдованныхъ законовъ роста, предположеніе, значительно уменьшающее вѣроятность конвергентнаго происхожденія. Послѣднее становится еще невѣроятнѣе, если припомнить, что въ структурѣ небно- челюстного аппарата мы имѣемъ дѣло не съ отдѣльно- выхваченнымъ признакомъ, не съ изолированной чертой строенія, а съ комплексомъ признаковъ, качественно отличнымъ отъ того, что наблюдается у неэгитогнатовъ, съ системой признаковъ, широко охватывающей цѣлую черепную область и сопровождаемой глубокимъ измѣненіемъ окружающихъ частей, короче — съ комбинаціей структурныхъ измѣненій, какъ нельзя лучше удовлетворяющей требованіямъ таксонамики въ томъ видѣ, какъ они формулированы Фюрбрингеромъ.
Таковы соображенія, побуждающія насъ, въ противоположность мнѣніямъ Gadow'а [20] и Newton'а [21] , высказаться противъ конвергентнаго развитія эгитогнатизма, принять для названнаго типа монофилетичное происхожденіе.
Что касается второго возраженія, а именно рискованности генеалогическихъ сопоставленiй, базируемыхъ исключительно на сходствѣ нѣкоторыхъ отдѣловъ черепа, то въ данномъ случаѣ, т.-е. при выведеніи родословной Passeres отъ Thinocorus'- образныхъ предковъ и разсмотрѣніи послѣдняго какъ ихъ видоизмѣненнаго потомка, названное возраженіе отпадаетъ по весьма простой причинѣ — отсутствію иныхъ моментовъ, которые позволили бы намъ связать организацію Passeriformes съ строеніемъ другихъ семействъ или отрядовъ. Въ самомъ дѣлѣ, если до Гексли напрасны были всѣ старанія орнитологовъ найти въ организаціи Passeriformes признакъ, общій всѣмъ представителямъ этой обширной группы, то не менѣе тщетными оказались всѣ дальнѣйшія попытки этого рода, предпринятыя послѣ появленія «Оn the Classification...» и единственной [22] чертой, дѣйствительно объединяющей всѣхъ Passeres, была и остается своеобразная архитектура небно-челюстного аппарата, причина, заставляющая насъ искать начало ихъ родословной вѣтви недалеко отъ того мѣста, гдѣ отвѣтвились примитивныя Limicolae [23].
Само собою разумѣется, что неблагопріятныя условія рѣшенія даннаго вопроса не устраняютъ принципіальной сущности указаннаго затрудненія, требованія — обосновать теоретически возможность установки генетической классификаціи на основаніи одного какого-либо признака. Говоря вообще, мы здѣсь имѣемъ дѣло съ чрезвычайно спорнымъ пунктомъ, съ кореннымъ логическимъ затрудненіемъ, неизбѣжно возникающимъ при переходѣ отъ понятія «морфологической непрерывности» къ понятію «непрерывности систематической» и связаннымъ съ необходимостью совмѣстнаго использованія признаковъ различныхъ категорій. Не касаясь вообще этого методологически довольно сложнаго вопроса — его детальное обсужденіе завело бы насъ слишкомъ далеко, — вернемся къ провизорно (т.-е. до нахожденія иного, болѣе надежнаго критерія) высказанному взгляду на значеніе эгитогнатизма, какъ признака, единственно пригоднаго для указанія предѣловъ и происхожденія цѣлаго отряда, и сопоставимъ это съ тѣмъ, что намъ извѣстно о таксономическомъ значеніи прочихъ типовъ.
Мы видѣли (см. Глава 1, Фактическія основанія классификаціи Гексли и позднѣйшая ихъ критика гл.), что съ точки зрѣнія діагностики принципы Гексліевой системы оказались мало продуктивными, и если бы въ установленіи естественныхъ подраздѣленій, путемъ анализа возможно большихъ признаковъ и различенія главнѣйшихъ типовъ измѣненій послѣднихъ, заключалась главная или единственная задача систематики, то, при доказанной преемственности всѣхъ четырехъ типовъ названной классификаціи и недіагностичности ея руководящаго критерія, можно было бы остановиться на приведенныхъ выше негативныхъ отзывахъ и удержаться вообще отъ пересмотра сочиненія, представляющаго только историческій интересъ. Въ действительности, какъ мы знаемъ, дѣло обстоитъ иначе: «Bei der genealogischen Reconstruction des natürlichen Systems, als des Stammbaums der Organismen, wird es daher nicht darauf ankommen, die einzelnen coordinirten und subordinirten Gruppen durch scharfe und exclusive Charakteristiken zu trennen, sondern vielmehr die vorwiegend erbliche oder angepasste Natur der Differential-Charaktere, ihr relatives Alter zu erkennen, und danach die gegenseitige Stellung der verwandten Gruppen zu bestimmen»... слова, которыми заканчивается одна изъ лучшихъ главъ классической работы Гэккеля [24] , съ которыми нельзя не согласиться. Но невозможно также не признать того, что въ противоположность первой, преимущественно разграничительной тенденціи, характеризующей младенческій періодъ въ исторіи классификаціи и вмѣстѣ съ тѣмъ начальную и неизбѣжную ступень какого ни на есть изслѣдованія, эта вторая часть работы, стремленіе генетически связать внутри изслѣдуемаго класса его различные отдѣлы или группы, независимо отъ ихъ объема и таксономическаго ранга, представляется гораздо болѣе трудной. Такъ, въ современныхъ орнитологическихъ системахъ естественность состава многихъ группъ не вызываетъ сильныхъ разногласій; что же до отношенія этихъ группъ между собою, ихъ относительнаго таксономическаго ранга и размѣщенія въ предѣлахъ категорій высшаго порядка, имѣющаго выразить частичный ходъ филогенеза, то можно безъ преувеличенія сказать, что съ появленія монументальнаго труда Фюрбрингера на родословномъ древѣ птицъ только немногія верхушечныя вѣтви подверглись спеціальной обработкѣ въ смыслѣ болѣе или менѣе точнаго опредѣленія ихъ относительнаго возраста и мѣста отхожденія, а въ остальномъ «а hundred classifiers, a hundred so called systems», и нѣкоторые авторы, довольствуясь характеристикой отдѣльныхъ категорій, воздерживаются совершенно отъ филогенетическихъ обобщеній за трудностью проблемы и недостаткомъ надлежащихъ признаковъ. Является вопросъ: какова природа этихъ признаковъ, необходимыхъ для возстановленія родословной, напр., для уясненія взаимныхъ отношеній основныхъ подраздѣленій класса, сравнительно немногихъ группъ, таксономически приблизительно равноцѣнныхъ? Очевидно, что то не могутъ быть черты «діагностичныя», такъ какъ онѣ суть выраженіе специфичности организаціи, результатъ морфологическаго обособленія, и соотвѣтственно тому являются для систематиковъ не столько орудіемъ синтеза, сколько средствомъ изоляціи и разграниченія. Это не могутъ быть также свойства, общія всѣмъ группамъ разбираемаго класса, ибо эти свойства говорятъ о существованіи одного общаго корня генеалогическаго древа и ни слова — о характерѣ его вѣтвленія. Несомнѣнно, что лишь такія особенности строенія, которыя представляются намъ въ формѣ болѣе или менѣе ясно выраженныхъ отличій, дающихъ непрерывный, связный рядъ послѣдовательныхъ измѣненій одного и того же признака и въ то же время выводимыхъ другъ отъ друга какъ различныя модификаціи одного или нѣсколькихъ типичныхъ состояній, сводимыхъ въ свою очередь къ однимъ исходнымъ отношеніямъ, — только такіе признаки пригодны для установленія филогенетическихъ связей. И въ этомъ смыслѣ qualitativ'ныя различія структуры небно-челюстнаго аппарата, какъ комбинаціи «морфологическихъ» особенностей черепа, охватывающихъ цѣлую скелетную область, даютъ намъ то, чего не въ состояніи были дать другіе признаки, какъ назовемъ здѣсь для примѣра, функціонально-важный, анатомически непостоянный фактъ развитія Carina Sterni и не менѣе спорадичные — ибо топографически не менѣе локализованные — признаки мускулатуры (Garrod) и системы органовъ кровеобращенія (Nitzsch). По крайней мѣрѣ, тѣмъ немногимъ, что извѣстно о координаціи главнѣйшихъ подотдѣловъ класса, мы преимущественно обязаны умѣлому и осторожному использованію принциповъ «Оn the Classification of birds», такъ, напр., признаніемъ примитивности Palaeognathae, ихъ полифилетичности, объединеніемъ отрядовъ Neognathae въ таксономическія единицы высшаго порядка — установленіемъ понятій Alecteromorphae, Pelargomorphae и Coraciomorphae.
Казалось бы, что этой положительной оцѣнкѣ признака противорѣчитъ явная искусственность нѣкоторыхъ типовъ; однако, неестественный составъ такъ наз. десмогнатовъ прекрасно согласуется съ новѣйшимъ [25] представленіемъ о десмогнатизмѣ, какъ понятіи, отнюдь не выражающемъ опредѣленнаго анатомическаго состоянія, но приложимаго къ структурамъ, морфологически глубоко различнымъ, хотя по виду и не всегда легко разграничимымь и имѣвшимъ каждая свою отдѣльную исторію. Напротивъ, что касается эгитогнатическаго типа, то на предшествующихъ страницахъ мы старались показать морфологическую цѣльность этого подраздѣленія и его вѣроятное монофилетичное происхожденіе. Такимъ образомъ, разсматриваемыя съ точки зрѣнія генеалогическаго синтеза, кажущіяся исключенія только подтверждаютъ положительное значеніе разбираемаго признака, доказывая лишній разъ, что въ погонѣ за этимологически-буквальнымъ примѣненіемъ термина изслѣдователи нерѣдко ищутъ и находятъ конвергенціи тамъ, гдѣ мы въ дѣйствительности имѣемъ дѣло съ явной, и порой довольно грубой аналогіей, и что за трудной и неблагодарной цѣлью — облечь разнообразіе видимыхъ структуръ въ дихотомически-абстрактную систему діагнозовъ — порой какъ будто забываются слова Фюрбрингера: «Verbinden, d. h. in der richtigen Weise Vereinigen ist mehr als Trennen».
─────── |
[7] «Son caractère semble d'abord purement négatif».
[8] Т.-е. Aegithognathismus, Parker (partim) & mihi, non «Aegithognathismus» Huxley & auct.
[9] Сверхъ того, дефектный въ отношеніи Proc. maxillo-palatini, утраченныхъ какъ и на объектѣ, послужившемъ описанію у Гексли. На одномъ скелетѣ, хранящемся въ музеѣ сравнительно-анатомическаго института московскаго университета, Proc. maxillo-palatini, судя по сохранившимся корневымъ отдѣламъ, по типу приближаются къ тому, что мы наблюдаемъ у другихъ эгитогнатовъ, и только вслѣдствіе чрезмѣрной тонкости срединнаго отдѣла, — области перехвата или шейки, — названные отростки легко утрачиваются во время препаровки.
[10] У Thamnophilus doliatus (loc. cit., I. p. 313).
[11] У Dendrocolaptes albicollis (p. 319).
[12] Говоримъ на основаніи аналогіи съ высшими эгитогнатами, за неимѣніемъ подъ рукою спиртового экземпляра черепа отъ Hemipodii.
[13] Такъ, напр., противорѣчивыя обозначенія хрящевыхъ участковъ, изображенныхъ между небными костями, прилежащими къ вершинѣ сошника: на рис. 2 означенные хрящи въ видѣ объемистаго парнаго образованія отмечены какъ «nasal fioor», тогда какъ на другомъ рисункѣ (3) мѣста ихъ занимаетъ пара гіалиновыхъ пластинокъ, названныхъ «interned alinasal wall».
[14] Возраженій Ньютона, имѣющихъ, въ своей оригинальной части, преимущественно теоретическій характеръ, мы коснемся ниже, при разсмотрѣніи методологическихъ сторонъ классификаціи.
[15] «Structure and Classification of Birds».
[16] Loc. cit., p. 37—39.
[17] Упоминается лишь вскользь, безъ надлежащаго рисунка...
[18] Pycraft. Some Points in the Morphology of the Palates of the Neognathae, p. 355.
[19] Мы имѣемъ здѣсь въ виду неубѣдительность и преждевременность такого «объясненія» для ряда частныхъ случаевъ, подобныхъ разбираемому примѣру, и не касаемся вообще теоретической стороны вопроса, одного изъ труднѣйшихъ въ современной біологіи.
[20] (2) р. 991.
[21] (12) р. 878.
[22] Если не считать особенностей въ расположеніи целомныхъ септъ (Beddard) и нѣкоторыхъ отличій въ прикрѣпленіи tensor m. patagialis brevis (Garrod) — признаковъ, мало пригодныхъ для классификаціонныхъ цѣлей и не строго характерныхъ...
[23] Ср. Fürbringer, р. 1234, 1315.
[24] Generelle Morphologie. Vierundzwanzigstes Capitel. VII. Charakter-Differenzen der subordinirten Gruppen, p. 402.
[25] Мы разумѣемъ группировку и характеристику различныхъ формъ «десмогнатизма», предложенныя Beddard’омъ (1), р. 151.