Более высокий уровень форм отражения характерен для многоклеточных животных, происшедших от одноклеточных организмов. У многоклеточных в связи с более сложными условиями существования возникла дифференцировка тела, появились ткани — покровная, сократительная, мускульная и проводящая возбуждение нервная ткань; образовались группы специальных, чувствительных клеток или рецепторов, воспринимающих из окружающей их внешней среды — раздражители разного качества (разной модальности) — химические, тепловые, механические, световые, звуковые.
У более высокоразвитых многоклеточных животных эти рецепторы преобразуются в сложные анализаторы (органы чувств), состоящие из воспринимающего прибора (рецептора), афферентных нервных волокон и соответствующих им нервных клеток — невронов, помещающихся в центральной нервной системе. Наличие сложных анализаторов, расширяя и углубляя ориентировку животного в окружающем мире, дает ему возможность не только воспринимать раздражения, идущие извне, как это осуществляется при посредстве рецепторов, но и анализировать восприятия, уточненно дифференцировать их модальность, их качество.
Органы чувств, как и специфичность их функций, также возникли в результате взаимодействия организма со средой, обусловившей их формирование, что обеспечивало организму более тонкое, точное и совершенное отражение внешнего мира[18] . К. Маркс определенно подчеркивал, что органы чувств являются продуктом всей истории развития живых организмов.
В процессе поступательного развития организмов, в процессе приспособления их к среде формирование органов чувств обеспечивало все более адекватное отражение раздражений, идущих из внешней и внутренней среды.
Развитие анализаторов теснейшим образом связано с развитием нервной системы и мозга, имеющим исключительно важное значение в прогрессивной эволюции многоклеточных животных.
Анализируя происхождение нервной системы в соотношении с другими тканями многоклеточных животных, А. А. Заварзин высказывает предположение, что нервная система возникла в тесной связи с мышечной системой, которая как бы «результирует деятельность нервной системы»[19].
В начальной стадии развития нервная система входит в состав единой примитивной нервно-мышечной системы, в рамках которой начинается дифференциация и нервной, и мышечной систем. Далее произошло разделение составляющих вначале единое целое функциональных компонентов, которые стали развиваться в виде соподчиненных друг другу нервной и мышечной систем. В доказательство этой гипотезы А. А. Заварзин приводит примеры из области развития этих систем в филогенезе организмов.
У одноклеточных организмов, по-видимому, нервные и мышечные компоненты объединены в так называемые мионемы, сократительные элементы, вызывающие ответные реакции инфузорий на раздражение. Аналогичная сократимость обнаруживается у примитивнейших многоклеточных животных — губок, еще не имеющих нервных элементов, но обладающих сократительными мышечными клетками, заключающими в себе, по-видимому, зачатки и тех признаков, которые характеризуют нервную систему.
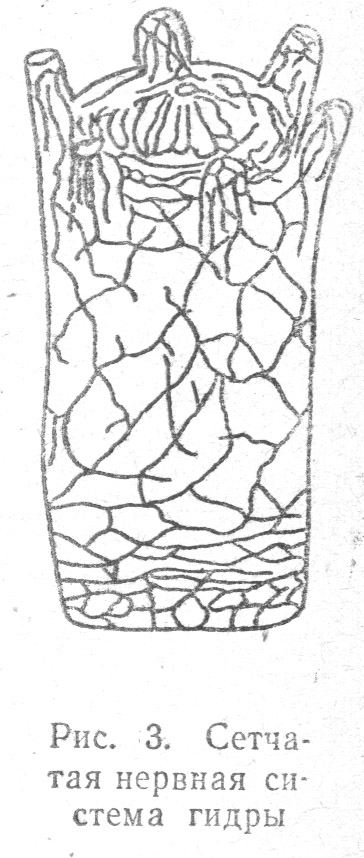
На следующем этапе филогенетического развития, как то наблюдается у кишечнополостных животных, имеется ясно выраженная сетчатого типа диффузная нервная система в виде рассеянных нервных клеток, пронизывающих все тело животного.
Нервная система кишечнополостных содержит два ряда дифференцированных нервных элементов: биполярные клетки с одним толстым коротким отростком, расположенные в наружном слое тела (гидры), и мультиполярные клетки с несколькими отростками, отходящие от базального конца биполярных клеток и погруженные более глубоко в промежуточном слое тела гидры, где они образуют сплетение и где помещаются мышечные клетки.
Следует допустить, что имеющиеся в наружном слое тела гидры биполярные клетки, и особенно их отростки, представляют собственно рецепторные элементы, а мультиполярные — являются передаточными элементами, переносящими раздражения на мышечные клетки.
Уже в сетчатой нервной системе кишечнополостных имеются все три элемента, необходимые для осуществления подобия рефлекторного акта[20].
Нервная система приняла на себя в организме роль интегрирующего аппарата, обеспечивающего по своей структуре и по своей функции организменную целостность и органическое единство живого существа[21].
Эволюция нервной системы в филогенезе организмов означала увеличение ее интеграционного значения для организма, а также увеличение ее взаимосвязи с различными его частями. Нервная система явилась основным посредником между организмом и окружающим миром, осуществляя в процессе рефлекторной деятельности взаимосвязь и взаимодействие среды и организма.
Как мы знаем, многочисленные рефлексы связывают живое существо как с внешней, так и с внутренней средой. Среда и организм, эволюционируя, находятся в теснейшей взаимной связи; организм изменяется под воздействием среды, приспособляясь к ней, что и обусловливает его сохранение и выживание.
«
Как часть природы, — пишет И. П. Павлов, — каждый
животный организм представляет собой сложную обособленную систему,
внутренние силы которой каждый момент, покуда она существует как таковая,
уравновешиваются с внешними силами окружающей среды.
Чем сложнее организм, тем тоньше, многочисленнее и разнообразнее элементы
уравновешивания. Для этого служат анализаторы и механизмы как постоянных,
так и временных связей, устанавливающие точнейшие соотношения между
мельчайшими элементами внешнего мира и тончайшими реакциями животного
организма. Таким образом, вся жизнь от простейших до сложнейших организмов,
включая, конечно, и человека, есть длинный ряд все усложняющихся до
высочайшей степени уравновешиваний внешней среды[22]
. Это уравновешивание осуществляется посредством нервной системы.
«
Свойства нервной системы животного более низкого уровня развития
трансформировались, переработались в свойства нервной системы более
высокого уровня развития. Эволюционный ряд современных животных это
— грубая копия того пути развития, который проделал животный мир в
течение многих тысячелетий.
В приблизительном виде сохранились общие черты, узловые пункты
эволюционного развития в виде классов, семейств, родов и видов, в свойствах
каждого из них отчетливо отображен свой собственный путь развития[23].
Изучение эволюции поведения представителей главных типов животного мира, выявление возрастания и усовершенствования ориентировки животных в окружающей их среде, как и освещение разных уровней развития их аналитико- синтетической деятельности, обеспечивающей организму тончайшие приспособления к непрестанно изменяющимся условиям существования, дает нам возможность конкретно обрисовать поступательный ход развития психики животных, уровня отражения ими внешнего мира в филогенезе организмов.
И. П. Павлов ясно подчеркивает, что «для того чтобы быть в равновесии с окружающим миром, надо, с одной стороны, как анализировать, так и синтезировать этот мир, потому что мир действует не только в виде простых агентов, но и в виде очень сложных комбинаций их, с другой — анализировать и синтезировать соответственно деятельность организма»[24].
Кишечнополостные — это весьма примитивные организмы, живущие в пресных или морских водах; тело их представляет двуслойный мешок. Наружная часть тела имеет защитные стрекательные клетки, а внутренняя — является пищеварительной полостью. Многие кишечнополостные (гидроидные полипы) ведут сидячий образ жизни, прикрепляясь нижним концом тела к почве; на верхнем конце, снабженном щупальцами, помещается ротовое отверстие полипов. Некоторые кишечнополостные (медузы), живущие в морях, превосходно плавают и весьма подвижны.
Кишечнополостные имеют ткани, части тела и органы. На этом этапе развития организмов у них появляется нервная система и их связи со средой становятся более многообразными и устойчивыми, чем у простейших.
Нервная система кишечнополостных еще весьма примитивна. Она состоит из нервных клеток с отростками, смыкающимися между собой в нервную сеть, пронизывающую все тело животного и особенно обильно разветвленную на щупальцах около рта; органах, широко вступающих в соприкосновение с предметами окружающего мира, в частности, с питательным материалом (Табл. 1.3). Первичная диффузная нервная система кишечнополостных не имеет центрального отдела: возбуждение, воспринимаемое в одном пункте тела животного, распространяется по всему организму.
Сетчатая нервная система кишечнополостных связана с мышечными элементами и находится с ними во взаимодействии, поэтому нервно-двигательные, «нейромоторные»[25] , реакции кишечнополостных, возникающие в любом месте тела в ответ на внешние воздействия, в зависимости от силы раздражителя, распространяются в большей или меньшей степени на все тело животного, вызывая его сокращение (например, у гидры).
Эти нейромоторные реакции являются прообразами рефлекторных реакций более высокоорганизованных беспозвоночных животных, имеющих централизованную нервную систему.
Кишечнополостные весьма чувствительны ко всякого рода раздражителям. Их чувствительные клетки, расположенные на всем теле, особенно густо приурочены к щупальцам — органам захватывания и притягивания пищи ко рту.
Рецепция кишечнополостных, определяющая проявление реакции питания, сводится к различению химических и осязательных раздражителей. При соприкосновении щупалец гидры с добычей (с маленькими рачками, которыми гидра питается) рецепторы щупалец возбуждают многочисленные мышечные клетки и стрекательные капсулы; далее раздражение передается через нервную сеть другим щупальцам и всему телу гидры; соседние щупальца стягиваются около захваченной добычи, обволакивают жертву, стрекательные капсулы опорожняются, обжигающая жидкость выбрасывается, парализует рачка; щупальца подносят рачка к ротовому отверстию гидры, последнее раскрывается и схватывает добычу.
Песок и несъедобные вещества гидра не поглощает. Это указывает, что нейромоторные реакции питания кишечнополостных приурочены к строго определенным раздражителям.
Кишечнополостные животные не ищут и не преследуют добычу, но захватывают ее при соприкосновении с ней. Деятельность нервной системы кишечнополостных ограничивается восприятием раздражений и согласованием работы отдельных частей тела (например, в акте питания).
Другой представитель кишечнополостных — актиния захватывает и заглатывает маленьких живых рыбок (Табл. 1.4), несъедобные вещества не вызывают у животного реакции заглатывания, а захваченные щупальцами — выбрасываются обратно; те же вещества, но пропитанные мясным соком, актиния поглощает.
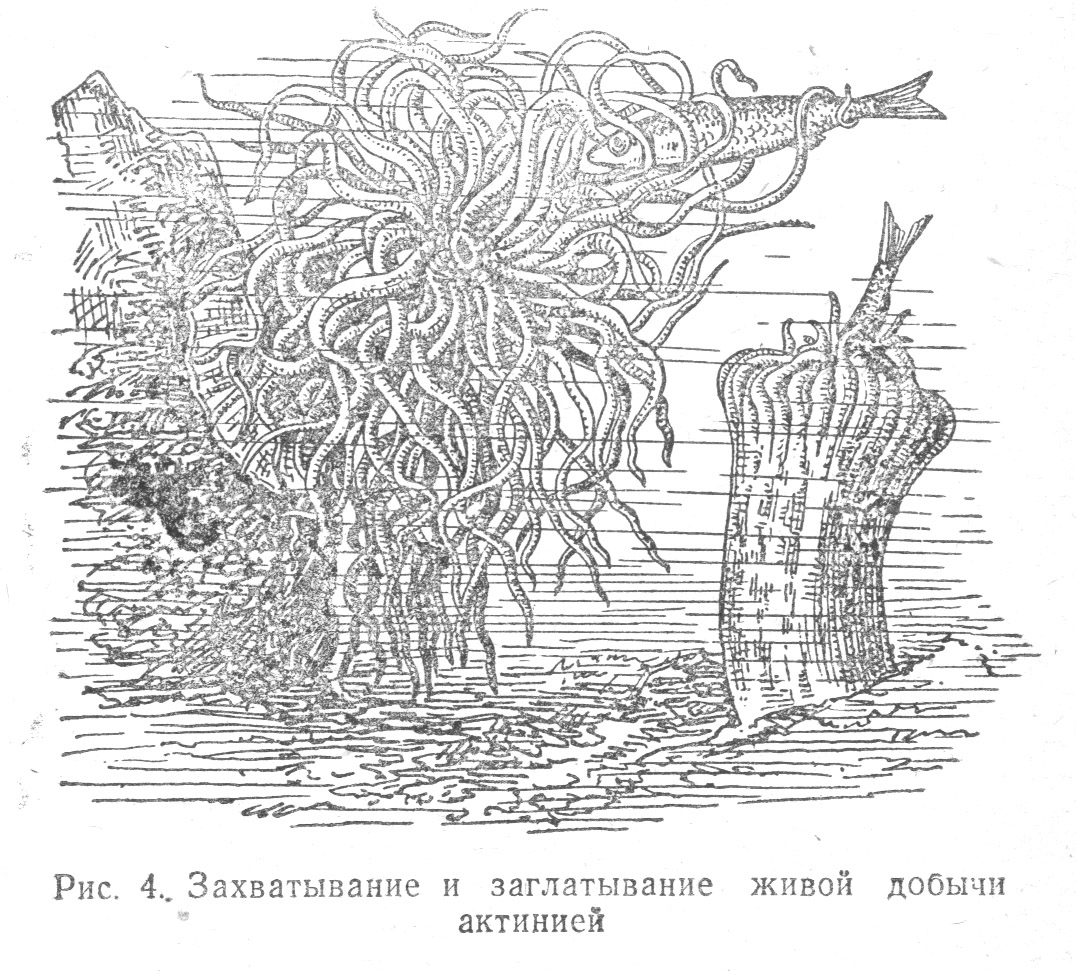
Замечательно, что и изолированные части тела (например, щупальца актинии) воспроизводят реакции, свойственные им, когда они составляют единое целое с животным.
Самостоятельность и адаптивность действий изолированных частей тела актинии объяснима автономностью отрезанных частей тела животного, заключающих в себе нервно-мышечные элементы. Отсутствие централизации нервной системы, генерализованный характер реакций и неразвитость тормозных процессов объясняют нам специфические особенности поведения кишечнополостных. Так, например, если актинии поднести на пинцете мясо и при захвате отвести его, не дав схватить, пустые щупальца все же будут сокращаться (несмотря на бесполезность этих действий) так же, как и соседние щупальца, получившие пищу.
У кишечнополостных наблюдаются нейромоторные реакции, относящиеся и к самозащите организма от вредящих воздействий. Например, при сильном механическом раздражении щупалец гидры, все тело животного съеживается в комочек и довольно долго остается в таком состоянии; организм воспроизводит простейшую форму защитной реакции. Но активной защиты у них нет, и, например, свободно плавающие медузы не могут реагировать своевременно на вредоносные воздействия среды или нападающего на них животного, уклоняясь от них.
Поведение кишечнополостных в основном обусловлено выработавшимися в процессе естественного отбора, прирожденными связями между определенными раздражителями и соответственными адаптивными реакциями организма на эти раздражители.
У кишечнополостных наблюдаются вариации поведения при повторяющихся или длительно воздействующих раздражениях и возможно возникновение адаптации в результате суммации раздражений: так при механическом сильном прикосновении и химических воздействиях у актиний вырабатываются временные связи как результат упражнения. Если, например, дать актинии фильтровальную бумагу впервые, то животное схватывает ее и проглатывает; позднее ту же бумагу актиния схватывает, но выбрасывает; через 2—5 повторений актиния отбрасывает бумагу прежде, чем последняя прикасается к ее рту.
Замечательно, что щупальца той же актинии, не участвовавшие в тренировке, не обладали этим опытом и брали фильтровальную бумагу даже тогда, когда она отбрасывалась натренированными щупальцами. Нескольких опытов тренировки и этих щупалец у того же животного было достаточно, чтобы и они реагировали так же, как и первая группа щупалец. Таким образом, опыт, полученный или испытанный одной частью тела, по нервной сети не передается другой части.
Согласно новейшим данным, у кишечнополостных (у гидры) удалось установить кратковременно сохраняющуюся временную связь, именно: если гидру, прикрепившуюся к краю водоема, ограничить в пространстве, т. е. дать ей возможность вытягивать щупальца и тело в воде лишь в одном направлении (например, влево), а потом постепенно, осторожным подливанием воды, создать такие условия, что гидра оказывается равномерно окруженной водой со всех сторон, то поведение ее меняется по сравнению с наблюдаемым в естественных условиях (Табл. 1.5).
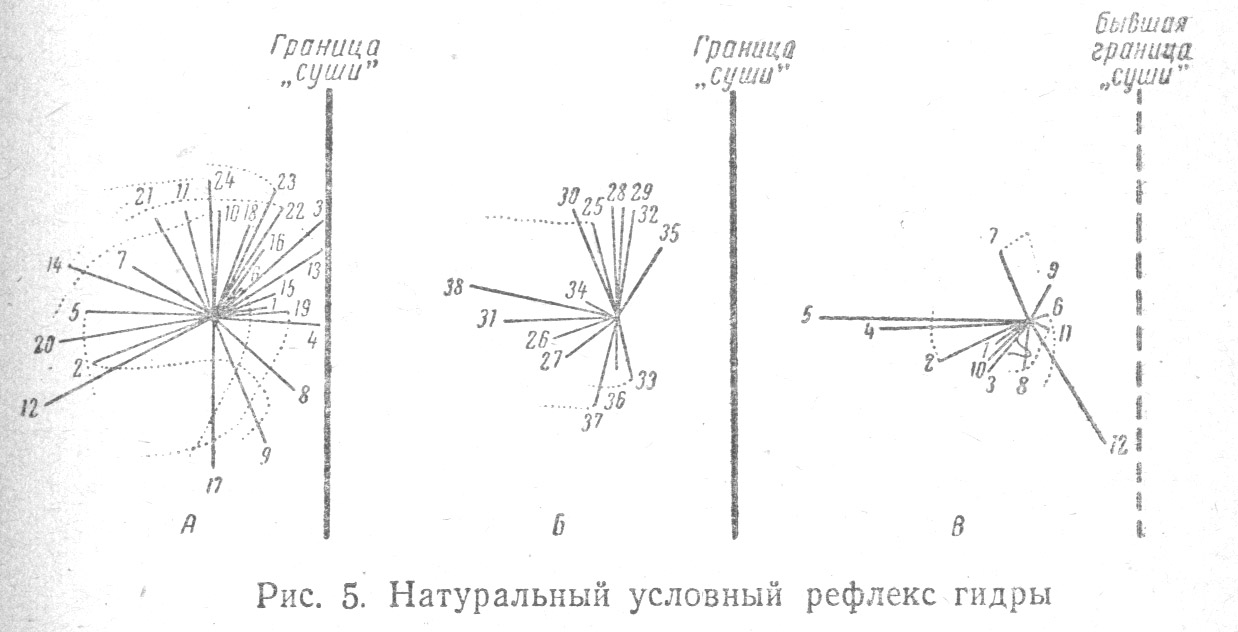
В то время как в обычных условиях, при обычном равномерном окружении гидры водой, движения ее щупалец бывают направлены равномерно в разные стороны, и количество этих движений во всех направлениях почти одинаково, то после ограничения движений гидры становится заметным преобладание движений щупалец в сторону свободного водного пространства. Выработанная временная связь неодинакового вытягивания щупалец (более частого и протяженного в левую и более редкого и короткого в правую сторону), сохраняется у гидры в течение часа и после ликвидации новых условий.
Только через три или четыре часа наблюдается угасание этой связи, и тогда гидра опять начинает производить щупальцами поисковые движения равномерно во все стороны[26].
У кишечнополостных совершенно не выражен инстинкт сохранения потомства, уход за молодью. Правда, у гидроидных живородящих полипов развивающаяся особь прикрепляется близ тела матери (где, по-видимому, находит больше пищи и защиты), но активного ухода родителей за своим потомством нет.
В итоге обзора поведения кишечнополостных мы должны сделать следующие выводы: организм этих животных реагирует на внешние воздействия, как целое, но у них отсутствует централизация нервной системы, их реакции носят генерализованный характер, процесс торможения еще не выражен.
Изменчивость поведения кишечнополостных весьма ограничена, а вызванное в условиях эксперимента изменение поведения не закрепляется. У кишечнополостных обнаруживается ярко выраженная парциальность реакций, проявляющаяся в автономности реагирования отдельных частей тела; опыт, полученный одной частью тела, не переносится на другую.
Изменение среды обитания, переход животных из водной среды в наземную и воздушную обусловили возникновение новых функций, связанных с изменением способов передвижения, строения тела, нервной системы и органов чувств. В соответствии с этим изменилось поведение животных, расширилась их деятельность и усложнились формы отражения ими окружающего мира.
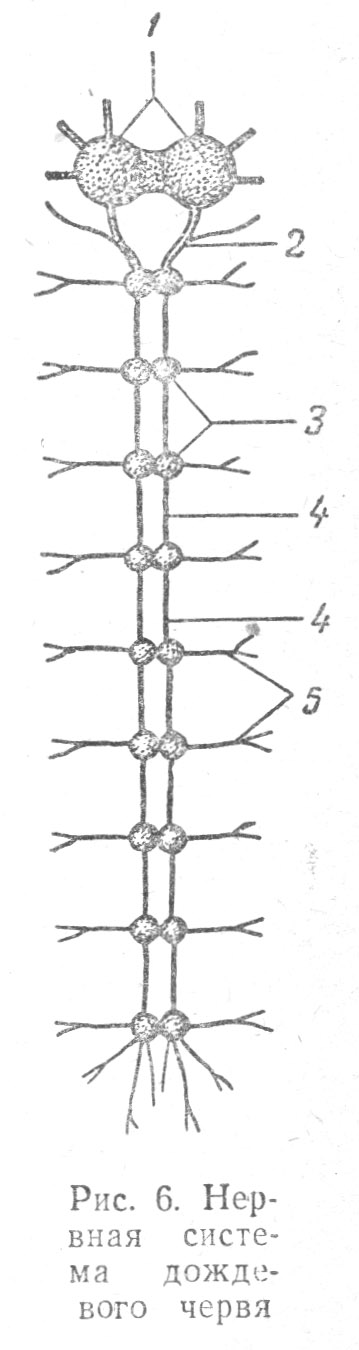
У червей, как ползающих животных, ведущих преимущественно роющий образ жизни, прокапывающих длинные норы в иле, земле или песке, выделяется ведущий головной отдел тела. Этот отдел первым встречается с новыми внешними воздействиями, поэтому на нем и возникают главные органы чувств: щетинки, усики, зачатки глаз. Сигналы от этих рецепторов поступают в головной ганглий, регулирующий поведение червя. На этой стадии развития животных возбуждение от рецепторов направляется по строго определенным нервным путям через центральные узлы и не распространяется во все части организма, как то наблюдалось у кишечнополостных.
Нервная система червей централизована (Табл. 1.6); в головном отделе червей образуются крупные узлы (или ганглии), от которых вдоль всего тела идут продольные нервные тяжи, соединяющиеся в каждом сегменте тела поперечно расположенными перемычками (или комиссурами). При расчлененности тела червя (у кольчатых червей) на сегменты меньшего размера, ганглии находятся и в каждом сегменте тела.
Отходящие от каждого ганглия нервы имеют чувствительные волокна, воспринимающие раздражения, и двигательные волокна, проводящие раздражения к мускулам или железам. Головные ганглии червей способны координировать их движения. Если, например, разрезать тело дождевого червя пополам, то каждая его половинка будет оживленно двигаться, так как заключает в себе ганглии, заведующие движениями. Но правильно, т. е. вперед, будет передвигаться лишь отрезанная передняя половина червя, содержащая головные ганглии, в то время как задняя половина вначале производит лишь бьющиеся, хаотические движения. Таким образом очевидно, что головной ганглий имеет особое значение по сравнению с другими. Если разрушить головные ганглии, то червь теряет нормальный образ действий: он уже не различает питательных и несъедобных предметов, при опасности лишь производит извивающиеся движения и долго не может успокоиться. Это также указывает на ведущую роль головных ганглиев червя. Преимущество головных (надглоточного и подглоточного) узлов перед другими узлами состоит в том, что они связаны с такими органами чувств, которые получают раздражения извне, не воспринимаемые другими частями тела червя.
Органы чувств низших плоских червей — щупальцы и аурикулярные органы с ресничками и щетинками, связанные с нервными клетками, могут воспринимать интенсивность химических и термических раздражителей.
У многощетинковых червей вся поверхность тела химически чувствительна.
Что касается органов зрения, то в группе червей мы находим все переходы их строения, начиная от стадии скопления пигментных клеток, переходя далее к стадии пузыреобразных глаз со светопреломляющими, светочувствительными элементами (у плоских и кольчатых червей) и, наконец, достигая этапа, когда имеются высокодифференцированные глазные камеры с роговицей, линзой, стекловидной массой, пигментными клетками (у многощетинковых трубчатых червей). У этих последних возможна даже аккомодация глаз, снабженных мускулами. Хотя различение формы предметов у червей не доказано, но определенно установлено, что черви могут различать свет, тень и направление света.
Плоские черви (планарии) отрицательно фототропичны: при различном освещении аквариума они скопляются обычно в затемненной его части (в естественных условиях они живут под камнями).
Более высокоорганизованные кольчатые черви (дождевые черви, живущие под землей) также негативно фототропичны. У них наиболее чувствителен к свету передний, головной конец тела, содержащий более густые скопления светочувствительных клеток, хотя эти последние разбросаны в коже и по всему телу. В меньшей степени светочувствителен задний конец червя и еще меньше средняя часть его тела в соответствии с неодинаковым распределением на этих частях светочувствительных клеток.
Наличие органов чувств (зачатков глаз; щупалец, щетинок, усиков) обусловливает то, что черви, в противоположность кишечнополостным, активно обеспечивают себя растительной пищей, например, дождевые черви схватывают двойные сосновые иглы за основание (где иголки скреплены), а упавшие листья за верхушки, за которые они втягивают их в свою норку (Табл. 1.7).
На основании этого явления одно время, вслед за Дарвином, ученые приписывали червям способность различать формы[27] , но контрольные опыты, произведенные Мангольдом[28] , показали, что черви различали в листе черенок от верхушки не по форме, но вследствие разного химического состава черенка и листовой пластинки, мякоти листа.
Таким образом, до сих пор не доказана способность червей к различению форм предметов.
У кольчатых червей играет роль осязательная и кинестетическая рецепция, связанная с их ползанием по земле, дающая восприятия контуров и пространственного расположения объектов окружающего их внешнего мира.

Под действием механических раздражителей (прикосновения) наземные кольчатые черви зарываются в землю, что и является их защитным приспособлением. Дождевые черви, вырывая себе в земле норки, прячутся в них при опасности; в норках они живут днем, туда же стаскивают пищу (опавшие с деревьев листья и сосновые иглы).
Морские реснитчатые черви во время прилива зарываются в песок, спасаясь от уноса их волнами с мест стоянок.
У червей иногда наблюдается при резких раздражениях быстрое и сильное сокращение передней части тела назад или изгибание в сторону, напоминающее движение «угрозы» более высокоорганизованных животных.
Для червей характерна безусловнорефлекторная деятельность, причем последняя может осуществляться не только целым животным, но и каждым отдельным сегментом его тела, который имеет свой ганглий — свой рефлекторный аппарат.
Например, обезглавленные черви, так же как и нормальные, будучи посажены на фильтровальную бумагу, в состоянии уползти с нее на землю, где они обычно пребывают.
Известный русский биолог-дарвинист В. А. Вагнер, специально занимавшийся экспериментальными исследованиями реакций отрезанных сегментов тела червей, пришел к выводу, что обезглавливание кольчатых червей (например, пиявок) не влечет за собой потери их способности к самостоятельным движениям, так как ганглии, заключенные в каждом сегменте их тела, делают этот сегмент до некоторой степени автономным носителем отдельных самостоятельно выполняемых функций[29].
У исходных форм кольчатых червей (многощетинковых червей) наблюдается инстинктивная безусловнорефлекторная строительная деятельность: они строят около своего тела трубки из клейких выделений своих желез и захваченных песчинок и камешков. Червь обволакивает этим клейким выделением желез песчинки и камешки и прикрепляет их к трубке, в которой сам помещается.
Некоторые черви этой группы (например, нереис — Nereis) в условиях эксперимента, даже будучи лишенными головных ганглиев, в состоянии производить своим передним концом типичные движения склеивания песчинок слизью, как только этот конец соприкоснется с песком.
У самки нереис есть зачатки проявления инстинкта ухода за потомством, выработавшегося в процессе эволюции сложной безусловнорефлекторной реакции, которая определенно приводит к охране развивающейся молоди. Именно: самка нереис после откладывания яиц и прикрепления их внутри своей жилой трубки начинает размахивать телом, нагнетая эмбрионам свежую воду, необходимую им для дыхания. Известны случаи, когда самка защищает свое потомство от приближающегося к ним самца, отгоняя его прочь.
У многощетинковых червей особи различных полов в период размножения активно ищут друг друга, осуществляя далекие странствования. Это связано во время созревания половых продуктов с действием взаимно возбуждающих химических раздражителей.
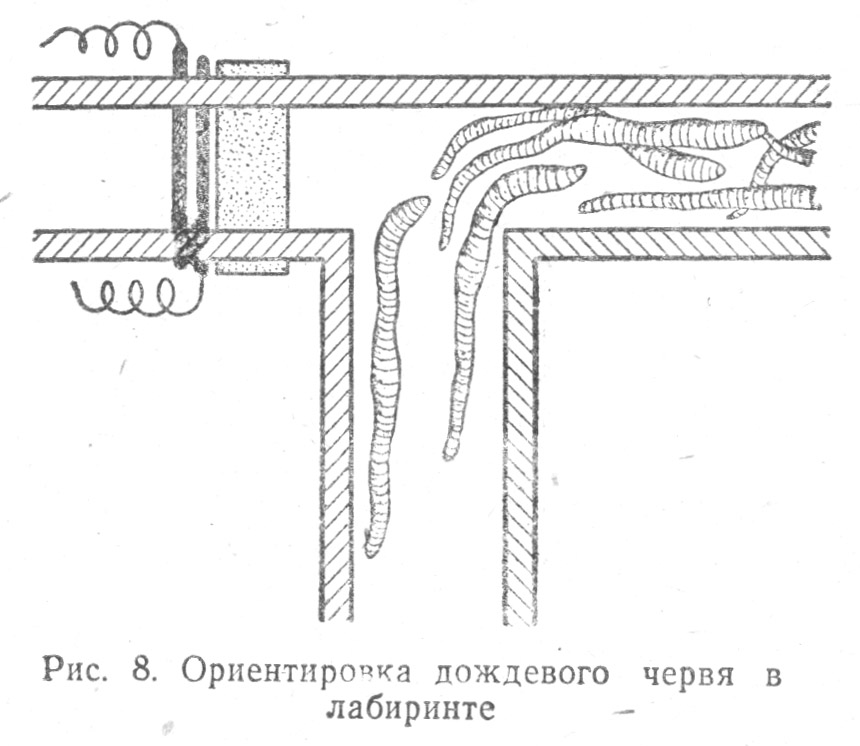
В условиях эксперимента у червей устанавливались условнорефлекторные связи: черви показали способность вырабатывать реакцию поворота в определенную сторону в Т-образном лабиринте (опыты Р. Иеркса)[30].
Если правая часть такого лабиринта имела выход и доступ к гнезду червя, а в левой червь получал удар электрическим током, то после 120—180 опытов проползания лабиринта червь приучался находить правильный путь, сразу продвигаясь в правое колено к гнезду (Табл. 1.8).
Если же после этого лабиринт перевертывали, т. е. в правом колене располагали провода под напряжением, а левое вело к гнезду, то червь был способен «переучиться», т. е. проползать в левый проход, ведший к гнезду, причем его переучивание происходило скорее, чем первое обучение (уже через 65 опытов у червя вырабатывался новый условный рефлекс).
Таким образом у червей обнаружили способность к установлению условнорефлекторных связей, и эти связи отличались у них известной пластичностью, их можно было переделать.
Замечательно, что новый навык проползания налево сохранялся и после того, как у червя были отрезаны шесть передних сегментов тела. Более того, даже лишенный этих сегментов, червь «выучивался», т. е. у него вырабатывался условный рефлекс правильной ориентировки в лабиринте, и нахождения пути к гнезду[31].
Условные связи удалось получить и у многощетинковых трубчатых червей.
Если червя (Nereis) при кормлении, когда он выползает из своей трубки, неизменно освещать, то через некоторое время он выползает из трубки и на одно освещение.
Сравнивая поведение червей с поведением кишечнополостных, мы обнаруживаем на этой фазе развития организмов ряд новых приобретений.
Наличие централизованной нервной системы обеспечивает большую активность животного в его взаимодействии со средой и переработку получаемых извне раздражений в головных узлах, хотя и наблюдается автономность реагирования отдельных сегментов тела, имеющих свои ганглии.
В связи с развитием многообразных органов чувств, расширяется отражательная способность червей.
В естественных условиях это выражается в осуществлении более сложных форм инстинктивного (безусловно-рефлекторного) поведения — в виде роющей, собирающей и строительной деятельности. В условиях эксперимента животное оказывается способным к изменению поведения, т. е. не только к выработке временных связей, но и к их переделке, осуществляющейся в более краткий срок, нежели первоначальное «выучивание». Это означает, что полученные осязательно-кинестетические раздражения оставляют следы в нервной системе животного и облегчают установление новых временных связей при пространственной ориентировке животного в новой ситуации.
Еще более высокий уровень развития форм отражения окружающего мы находим у членистоногих животных, нервная система, органы чувств и рецепция которых, по сравнению с червями, чрезвычайно осложнились в связи с высокой степенью приспособления этих животных к разнообразнейшим условиям существования: к жизни в воде, на земле и в воздухе.
Членистоногие животные включают несколько классов: ракообразных, паукообразных и насекомых.
Особенности строения тела членистоногих состоят в наличии у них плотного наружного хитинового покрова, имеющего опорную и в то же время защитную роль. Тело членистоногих разделено на 2—3 части: голову, грудь и брюшко, или головогрудь и брюшко. Тело, как и конечности, членистоногих состоит из отдельных члеников. Строение их конечностей весьма разнообразно в связи с приспособлением к ползанию или беганью по земле, плаванию в воде, прыганью и летанию в воздухе.
Образ жизни членистоногих, способы добывания ими пищи, как и характер пищи, чрезвычайно разнообразны у представителей разных классов. Нервная система членистоногих состоит из надглоточного ганглия, окологлоточных комиссур и брюшной нервной цепочки, проходящей вдоль всего тела, имеющей в каждом сегменте небольшие ганглии.
Головной ганглий членистоногих не только является координирующим центром, но оказывает задерживающее влияние на протекание безусловных рефлексов. Например, при разрезании (у ракообразных) связей между головным ганглием и брюшными стволами оперированный краб схватывает ротовыми частями и перерабатывает предложенную ему пищу в таком количестве, что битком набивает желудок, и последний лопается, так что краб погибает[32].
У наиболее высокоорганизованных членистоногих — насекомых — нервная система часто осложняется благодаря слиянию ганглиев, приуроченных к голове, груди, брюшку, в результате образуются головные, грудные и брюшные узлы. Особенное развитие получает головной узел, связанный с органами чувств: зрением, обонянием, осязанием (Табл. 1.9).
Передняя часть надглоточного ганглия насекомых является зрительной частью, от нее идут нервы, направляющиеся к глазам, задняя часть — обонятельная, от средней части переднего ганглия идут нервы к усикам, а также отходит окологлоточная комиссура, связывающая надглоточный ганглий с подглоточным ганглием, который иннервирует роговые части и находящиеся на них вкусовые органы. Особенно важной частью переднего отдела головного ганглия насекомых являются «грибовидные», или «стебельчатые» тела; по-видимому это важные ассоциационные центры, особенно хорошо развитые у колониальных насекомых (муравьев, пчел, ос, термитов), обладающих сложными инстинктами.
У полиморфных насекомых (пчел, муравьев), т. е. разделяющихся на самцов, рабочих и маток — плодущих самок, — замечается наибольшее развитие грибовидных тел у рабочих, меньшее — у маток и еще меньшее, почти рудиментарное состояние их у самцов.
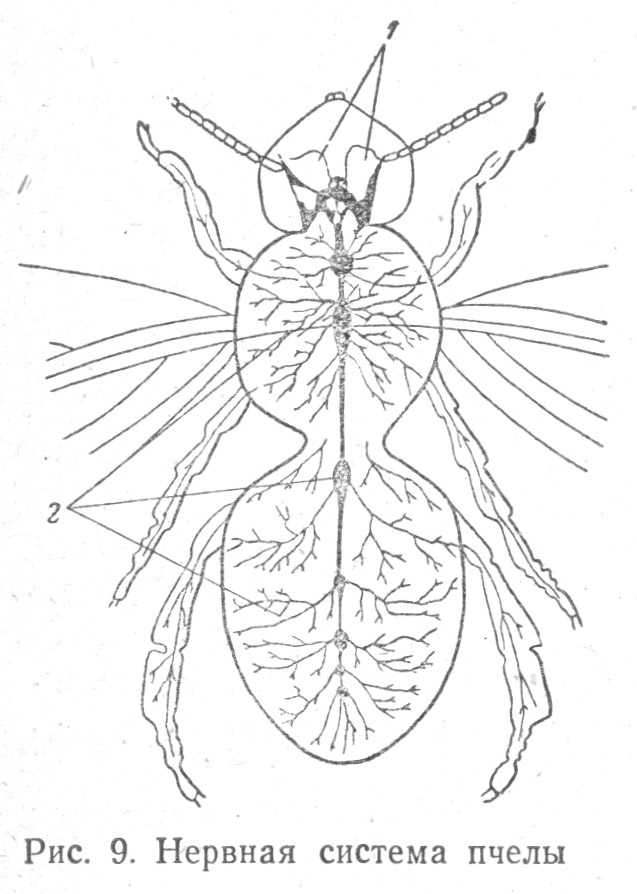
Установлено, что при переходе одиночных перепончатокрылых насекомых к жизни в сообществе, т. е. при осложнении условий их существования, усиливается развитие их грибовидных тел.
У насекомых имеются разнообразные рецепторы весьма сложного строения. У них хеморецепция дифференцирована на обонятельную и вкусовую. У представителей ранее рассмотренных типов беспозвоночных (кишечнополостных и червей) обоняние и вкус слиты.
Вкусовые рецепторы, или густорецепторы насекомых, приурочены главным образом ко рту, но у некоторых групп мух и у бабочек вкусовоспринимающими поверхностями являются кончики лапок.
Густорецепторы лапок, прикоснувшихся к вкусовым предметам, дают сигналы, которые позволяют легко дифференцировать, например, сладкую жидкость, от неподслащенной воды, однако порог различения сладкого у мух в 20 раз ниже, чем у человека. Насекомые различают четыре вкусовых качества: сладкое, горькое, соленое и кислое.
Обонятельные рецепторы насекомых расположены на усиках, и если, например, отрезать усики у тараканов, то животные уже не могут находить подброшенную им пищу и погибают. Обонятельные рецепторы, как это доказано опытами Фриша, хорошо развиты и у пчел.
Фриш сочетал посещение пчелами коробок, где помещалась подсахаренная вода, с дополнительным раздражителем — запахом эфирного масла: при этом оказалось, что в последующем пчелы летали в коробки только на запах, хотя поощряющая их пища не вкладывалась. На основании этих опытов Фриш пришел к заключению, что пчелы способны различать такие же разнообразные запахи, как и человек[33].
У муравьев обоняние, как и зрение, играет большую роль. Обоняние помогает муравьям ориентироваться в пространстве при нахождении пути к гнезду после выхода из него. Когда же они теряют обонятельные следы, то пользуются зрением, руководствуясь направлением солнечных лучей.
Хеморецепция — именно запах — играет большую роль, особенно у колониальных насекомых — пчел при опознавании чужих особей. Если, например, в один из ульев, не имеющих пчелы-матки, подсадить матку из другого улья, то пчелы ее тотчас же убивают; если же пересаживаемую пчелу предварительно продержать в улье пчел-хозяев, изолировав в маленькой клеточке, где она примет запах гнезда хозяев, то чужая пчела может быть посажена в гнездо безнаказанно.
Дифференцирование по запаху своих от чужих характерно и для муравьев. Чужака-муравья, попавшего в гнездо муравьев-хозяев, после ощупывания его усиками, обычно изгоняют или убивают. Если же враждующим муравьям из разных семей отрезать усики, они мирно сожительствуют в одном гнезде. Хеморецепция насекомых играет большую роль и при распознавании полов. По запаху бабочки-самцы находят бабочку-самку даже в том случае, если последнюю посадить в коробку.
Некоторые бабочки-самки (например, тутового шелкопряда) особенно пахучи, и оказалось, что к одной из таких бабочек, посаженной в коробку на 6,5 часов, слетелось 127 бабочек-самцов.
Хеморецепторы и у бабочек находятся на усиках, и если самцам бабочек отрезать усики, то они не находят самку.
Обоняние и осязание насекомых связано с особой так называемой топохимической чувствительностью.
Осязательные органы чувств, или тангорецепторы, у членистоногих находятся в усиках, антеннах и щупальцах.
У насекомых, в частности у тараканов, усики играют не только роль хеморецепторов, но и тангорецепторов: тараканы с отрезанными усиками не в состоянии правильно ориентироваться в пространстве.
Органы зрения у насекомых очень развиты: это сложные фасеточные глаза, состоящие из множества мелких глазков, или омматидиев. Подобные глаза дают мозаичное изображение, которое является комбинацией множества мелких частичных изображений, сливающихся и дающих единое прямое изображение предмета.
Хотя у насекомых глаза хорошо развиты, но они отчетливо различают лишь маленькие и близкие предметы, в то время как большие и далекие видят неясно.
У насекомых имеются не только сложные фасеточные боковые глаза, но у некоторых также и глазки: первые воспринимают расположение предметов, а последние позволяют ориентироваться в отношении к горизонту и действуют при слабом свете.
Глазки насекомых имеют большой крупный хрусталик, пропускающий большое количество лучей; полагают, что они приспособлены для видения при слабом свете, например, в ульях и в муравьиных гнездах.
Быть может, глазки содействуют определению расстояния при ориентации насекомого в пространстве и при различении расстояния между предметами.
Экспериментально установлена способность насекомых реагировать на движение предметов с расстояния от 1,5 до 2 м. По-видимому, насекомые в состоянии различать мельчайшие детали предметов, невидимые для человеческого глаза, но лишь на очень близком расстоянии (1 м от глаза); на большом расстоянии фасеточные глаза дают неясное изображение.
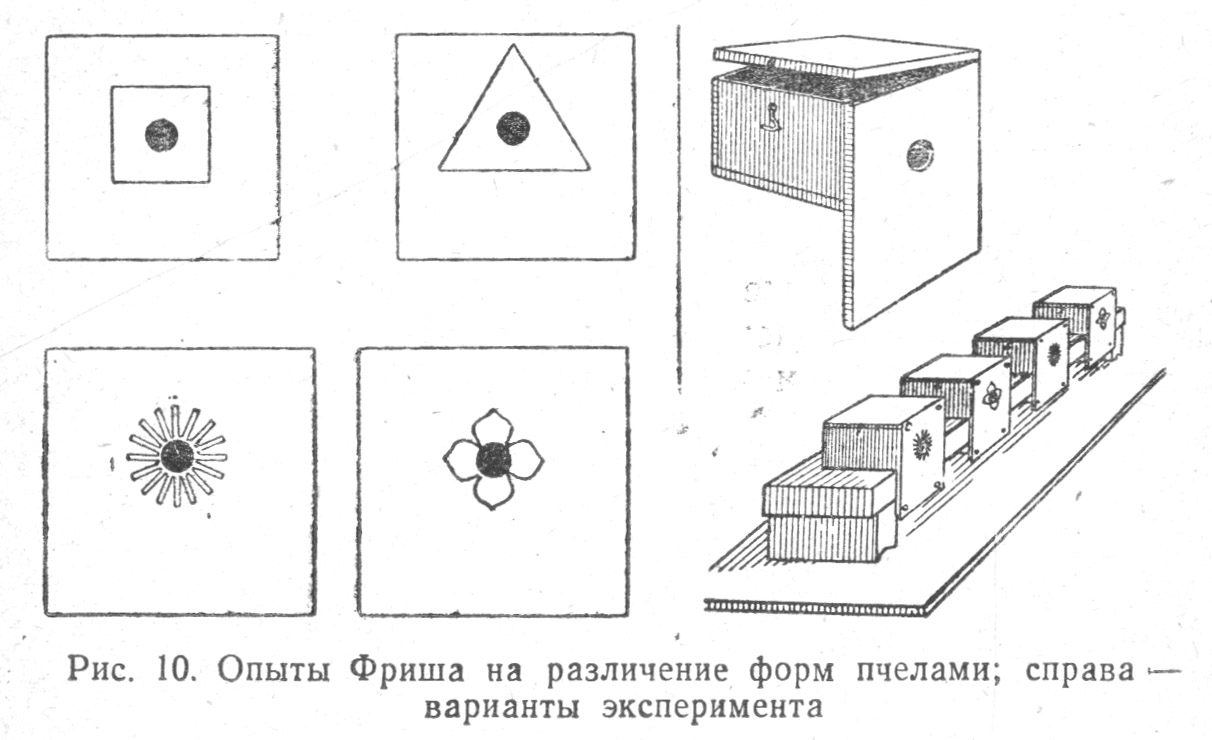
В экспериментальных условиях определенно доказана способность насекомых, в частности пчел, различать цвета. Границы цветного зрения пчел находятся между цветами с длиной волн 650—300 мμ.
Но спектр, видимый пчелами, укорочен в красной части (по сравнению с таковым человека) и удлинен в сине-фиолетовой. Доказано, что пчелы различают четыре цвета: желтый, сине-зеленый, синий и ультрафиолетовый. Этот последний цвет не воспринимается человеческим глазом, а пчелы воспринимают его как самостоятельный цвет (муравьи также способны различать этот цвет). Но пчелы слепы на красный цвет, который различают бабочки, и если пчелы и посещают красные цветы, например мака, то этот цветок имеет для них ультрафиолетовую окраску. Цветы многих европейских растений, которые мы называем красными, на самом деле имеют пурпурную окраску и пчелам кажутся синими[34] . Близкие цвета одной и той же половины спектра пчелы нередко смешивают, например, желтый и оранжевый с зеленым, синий — с фиолетовым и пурпурным. Особенно хорошо удавалось дрессировать пчел на голубой и синий цвета. Пчелы явно предпочитают при свободном выборе синий и пурпурный цвета. К тому же выводу еще раньше Фриша пришел и замечательный исследователь насекомых Джон Леббок[35].
Признак цвета имеет для пчел явное биологическое сигнальное значение.
Наблюдение Фриша над пчелами показало, что при посещении цветов они прежде всего ориентируются по зрению, и только на ближайшем расстоянии от цветов — по запаху[36].
Пчелы различают и форму предметов; они могут распознавать определенный рисунок венчика цветка, имеющего форму обычно посещаемых пчелами цветов (Табл. 1.10).
Пчелы лучше различают очертания рисунков, сходных с таковыми биологически значимых предметов (венчиков цветов), нежели биологически нейтральных — например, геометрических форм (квадратов и треугольников)[37].
Таким образом, обнаруживается, что признаки, имеющие для животных сигнальное значение, распознаются ими лучше, чем признаки биологически нейтральные, т. е. не столько строение глаза и его способность видения, сколько сигнальность признаков определяет, что именно из окружающего воспринимается насекомым. Доказано реагирование на форму предметов у стрекоз, у хищных мух, пчел, муравьев. Положительные фотокинетические реакции насекомых некоторых отрядов обусловливают то, что при наличии искусственных источников света мы видим массовый полет к ним и массовую гибель насекомых (например, бабочек, мух).
Пчелы могут различать определенные предметы, служащие им вехами при нахождении гнезд, и отыскивают последние по этим вехам. Так, например, при перестановке улья на 2 м с востока на запад пчелы собирались в воздухе в том месте, где было прежде летное отверстие. В течение 5 минут они кружились у этого места, а потом полетели к новому местоположению улья.
Но если пчелам отрезали усики, они тотчас же подлетали к переставленному улью. Это означало, что в первом случае они ориентировались в пространстве преимущественно по кинестетическим сигналам, после удаления усиков они стали ориентироваться по зрению[38] . В. А. Вагнер убедительно доказал экспериментами, что шмели при розыске нектароносных растений, при собирании нектара руководствуются исключительно зрением[39].
Звуковые волны воспринимаются у животных фонорецепторами, появляющимися в филогенезе организмов значительно позднее других рецепторов. Они обнаруживаются именно у членистоногих в виде тимпанальных органов, помещающихся у насекомых в разных местах тела и состоящих из тонкой хитиновой оболочки — барабанной перепонки, тимпанум, к которой прилегает как бы резонатор трахейный ствол голени (у кузнечика).
Обычно тимпанальные органы имеются у тех насекомых, которые сами издают звуки трением крыла о крыло или о лапку, например, у сверчков и кузнечиков. Воспроизведение звуков играет большую роль в нахождении самцами самок — в привлечении друг к другу разных полов.
Анализаторная способность членистоногих, связанная с дифференцированием самых разнообразных свойств биологически значимых предметов, воспринимаемых при помощи органов обоняния, вкуса, зрения, осязания и слуха, сочетается с высокой степенью развития синтетических процессов, проявляющихся в осуществлении активных связей животного не только с предметами окружающей среды, но и с особями своего вида, в самых разнообразных инстинктивных формах их поведения.
У членистоногих мы обнаруживаем чрезвычайно сложную инстинктивную деятельность, наступающую в ответ на определенные раздражители.
И. П. Павлов определяет инстинкты как сложнейшую безусловнорефлекторную деятельность, давая им следующую характеристику: «К этим обычным рефлексам, давнему объекту исследования физиологов в их лабораториях и касающимся главным образом деятельности отдельных органов, примыкают еще прирожденные реакции, также при посредстве нервной системы происходящие закономерно, т. е. строго приуроченные к определенным условиям. Это реакции разнообразных животных, касающиеся деятельности целого организма под видом общего поведения животных и обозначаемые особым термином „инстинктов“»[40].
Инстинкты насекомых по сравнению с таковыми других животных отличаются особой сложностью. Именно в этой группе организмов развитие инстинктов достигает весьма высокой степени. Они приобретают цепной характер, включая длинные серии последовательно и преемственно развивающихся действий приспособительного характера.
Механизм их осуществления заключается в том, что в этих цепных рефлексах конец одного рефлекса есть возбудитель следующего[41].
Проявление инстинктов зависит не только от наличия определенных внешних стимулов, но и от внутреннего физиологического состояния организма, деятельности желез внутренней секреции (гормональных факторов) и гуморальных влияний, связанных с состоянием крови.
Инстинктивная деятельность насекомых является преобладающей формой их поведения, при которой организм уравновешивается со средой. По этому поводу И. П. Павлов говорит следующее: «Как определенная замкнутая вещественная система он (т. е. организм — Н. Л.- К.) может существовать только до тех пор, пока он каждый момент уравновешивается с окружающими условиями. Как только это уравновешивание серьезно нарушается, он перестает существовать как данная система. Рефлексы суть элементы этого постоянного приспособления или постоянного уравновешивания»[42] и далее. «Совокупность этих рефлексов составляет основной фонд нервной деятельности как человека, так и животных»[43].
Сложнейшие рефлекторные реакции животных, лежащие в основе их инстинктивного поведения, направлены на осуществление какой-либо биологически значимой для животного цели, удовлетворяющей его потребности.
Фактически в конкретном поведении животного мы имеем сложное переплетение безусловных и условных рефлексов. «В чистом виде безусловная реакция проявляется один лишь раз при рождении организма; в дальнейшем эта реакция вступает в контакт с компонентами среды, включаясь в сложную систему средовых отношений»[44] . Таким образом, в последующей жизни животного безусловные и условные рефлексы переплетаются в единой рефлекторной деятельности настолько тесно, что их трудно расчленить.
Поведение каждого животного приспособительно: чтобы выжить на жизненной арене животное должно приспособиться к условиям своего существования.
Инстинктивные формы поведения целесообразны лишь при узко ограниченных условиях их проявления, они становятся нецелесообразными при резком отклонении от этих условий.
Инстинктивные формы деятельности сравнительно стереотипно протекают у всех особей одного вида животных. В основном наблюдается целесообразность их осуществления без предварительного обучения, но все же сложная, например, строительная инстинктивная деятельность молодых животных, впервые приступающих к гнездостроению, осуществляется несколько хуже, чем старых. Это указывает, какое большое значение имеет наслоение условных рефлексов на основной фонд врожденных, безусловных.
Сложнейшие инстинктивные формы поведения членистоногих особенно ярко выражены у паукообразных и у насекомых, у которых нервная система имеет значительно более сложное строение, чем у представителей выше рассмотренных типов животных (кишечнополостных и червей).
Неудивительно, что членистоногие обладают весьма сложным поведением, относящимся к основным проявлениям их жизни — актам питания, самосохранения и размножения.
В проявлении различных инстинктивных форм поведения членистоногих мы наблюдаем расширение пределов ориентировочной деятельности и связанных с ней инстинктивных реакций. Это последнее особенно ярко сказывается в проявлении инстинкта питания и инстинкта ухода за потомством.

Так, если у кишечнополостных животных мы отмечали проявление акта питания лишь при случайной встрече с пищей, если черви (дождевой червь) уже не только осуществляют поиски пищи, но и активно выбирают ее и втягивают в норку, собирают и сохраняют, то у членистоногих, например, у некоторых пауков, мы встречаем сложные приемы деятельности, предваряющие акт питания, связанные с устройством специальных «ловушек» для захватывания живой жертвы.
Напомним классические примеры, относящиеся к устройству тенет пауками (Табл. 1.11). У паукообразных имеются паутинные железы, которые выделяют клейкое, быстро твердеющее на воздухе вещество, дающее шелковистую паутинную нить. Из этих нитей пауки и вьют тенета-ловушки для живой добычи. Из паутинных нитей пауки делают кокон для помещения в него своих яиц и выстилают ими внутреннюю поверхность своих норок.
У паукообразных превосходно развиты осязательные рецепторы (тангорецепторы), и это имеет большое значение для пауков, ловящих добычу с помощью тенет. Соткав искусную ловушку, растянутую между несколькими пунктами опоры, сам паук обычно помещается где-нибудь поблизости в особом укрытии — гнезде. К этому гнезду от центра паутины идет особая соединительная «сигнальная» нить, которую паук, сидящий в гнезде, держит двумя передними лапками. Если какое-нибудь насекомое залетит и застрянет в паутине, паутина сотрясается, ее колебания немедленно передаются по «сигнальной» нити пауку, он выбегает к паутине, кусает челюстями жертву, запутывает ее выделяемой паутиной и в запакованном виде уносит в гнездо, где ее и съедает. Если паук очень сыт, он оставляет добычу в паутине.
Замечательно, что паук реагирует лишь на паутинные колебания определенной силы и амплитуды, т. е. на адекватные раздражители, возникающие при движениях в паутине обычных его живых жертв (мух и других небольших насекомых), на другие колебания паутины он не реагирует. Еще более интересно, что паук, даже голодный, встретив муху вне паутины, убегает от нее, так как он воспринимает муху как добычу лишь при определенных условиях — при нахождении ее в паутине, т. е. в строго определенной обстановке.
Это говорит о ситуационной связанности восприятий предметов у животных на первых этапах предметного восприятия (как это имеет место в приведенном примере у пауков).
Еще более сложна инстинктивная деятельность, связанная с ловлей живой добычи, у одного замечательного насекомого — личинки муравьиного льва.
В статье «Муравьиные львы и их ловчие воронки» Пузанова- Малышева сообщает, что эта личинка, забравшись в песок, движется по его поверхности, оставляя круглую борозду, при этом она продвигается задом вперед, часто по часовой стрелке. Слегка зарывшись в песок, личинка набирает его на свое тело и ритмическими движениями головы сбрасывает песок в сторону за линию круга; далее личинка делает второй круг и третий, все меньшего и меньшего диаметра. Эти круговые хождения длятся до тех пор, пока в центре круга не образуется ямка в виде правильной воронки[45] (Табл. 1.12). Как только на край воронки попадает какое-нибудь насекомое, личинка взметывает снизу фонтанчик песка, которым и сбивает внутрь воронки попавшую в западню жертву. Если насекомое не удается сбить с одного раза, личинка выбрасывает новые порции песка до тех пор, пока жертва не свалится внутрь воронки. Тогда личинка схватывает добычу своими сильными челюстями, погружает в песок, потом треплет и высасывает. Кусая жертву, личинка впускает в ранку каплю яда, который парализует насекомое.

Здесь мы обнаруживаем сложную, стройно смыкающуюся цепь безусловнорефлекторных актов, связанных с питанием личинки.
Некоторые авторы (Doflein)[46] , исследовавшие поведение муравьиного льва, делали вывод, что муравьиный лев является «рефлекторным автоматом», т. е. что его поведение при ловле добычи есть ряд механически наступающих реакций на раздражители среды.
Но ближайший специальный анализ этого сложного инстинктивного поведения, произведенный Пузановой-Малышевой, обнаружил, что проявление этого инстинкта не абсолютно стереотипно: при изменяющихся условиях организм может в соответствии с новой обстановкой до некоторой степени изменить свое поведение[47] . Это указывает, что инстинктивное поведение в известной мере пластично, оно связано с образованием условных рефлексов, с использованием насекомым своего индивидуального опыта и приспособлением его к достижению биологически значимой цели[48].
У хищных насекомых мы наблюдаем преследование живой добычи, ее поимку, умерщвление и использование для питания будущих личинок; у перепончатокрылых — далекие полеты за пищей, сбор нектара, его сохранение и запасание для сочленов улья и питания личинок.
Не менее сложна инстинктивная деятельность ракообразных и насекомых, направленная на достижение другой биологически значимой цели, например, самозащиты.
Замечателен, например, сложный инстинкт маскировки у крабов, выражающийся в том, что краб схватывает и держит над своей головой мертвых крабов, головы мертвых рыб и другие предметы, служащие ему щитами от врагов. В этом случае мы имеем зачаток опосредствованных действий, когда животное употребляет для защиты не свой собственный орган, например, клешню, но берет предметы из окружающей среды и использует их для самозащиты.
Крабы, обладающие шипами на ногах и спине, насаживают на эти шипы зеленые водоросли, превосходно скрывающие их на зеленом фоне растений. Еще замечательнее у крабов опосредствованное употребление предметов, подвергшихся предварительной обработке. Оторвав небольшой кусок водорослей, краб обрабатывает его челюстями (или ногами) до тех пор, пока тот не скатается в клубок, который краб схватывает своими длинными клешнями и кладет на шипы, торчащие на его голове и спине, закрепляя водоросли движением ноги и не прекращая своей деятельности до тех пор, пока не покроет все свои свободные шипы.
В условиях эксперимента было установлено, что крабы как бы маскировались в те цвета, которые соответствовали фону среды: в зеленом аквариуме они выбирали из красных и зеленых бумаг — зеленые, в красном — маскировались в красные, отличая их от белых (но крабы не различали желтых и зеленых цветов).
Экспериментальный анализ этого хроматического миметизма у крабов, произведенный Минкевичем[49] , привел к заключению, что это явление объясняется образованием зрительных рефлексов на те или другие цвета.
Поведение животного после длительного пребывания в среде определенной окраски меняется, и животное начинает реагировать лишь на эту окраску и негативно на всякую другую.
Контрольный опыт содержания крабов сначала в одноцветных аквариумах (одних в красном, других — в зеленом), а затем в двухцветном (наполовину красном, наполовину зеленом) обнаружил, что крабы пошли в ту часть аквариума, которая соответствовала цвету прежней их обстановки. Однако крабы, содержащиеся в аквариуме красного цвета и замаскированные красными бумажками, при пересадке их в аквариум другого цвета, например, зеленый, бумажки уже не меняли, хотя оказывались, заметными на окружающем фоне. Это указываю, что хотя в основном явление описанного приспособительного миметизма имело инстинктивный характер, но в нем проявлялись и условнорефлекторные наслоения, связанные со способностью крабов различать некоторые цвета.
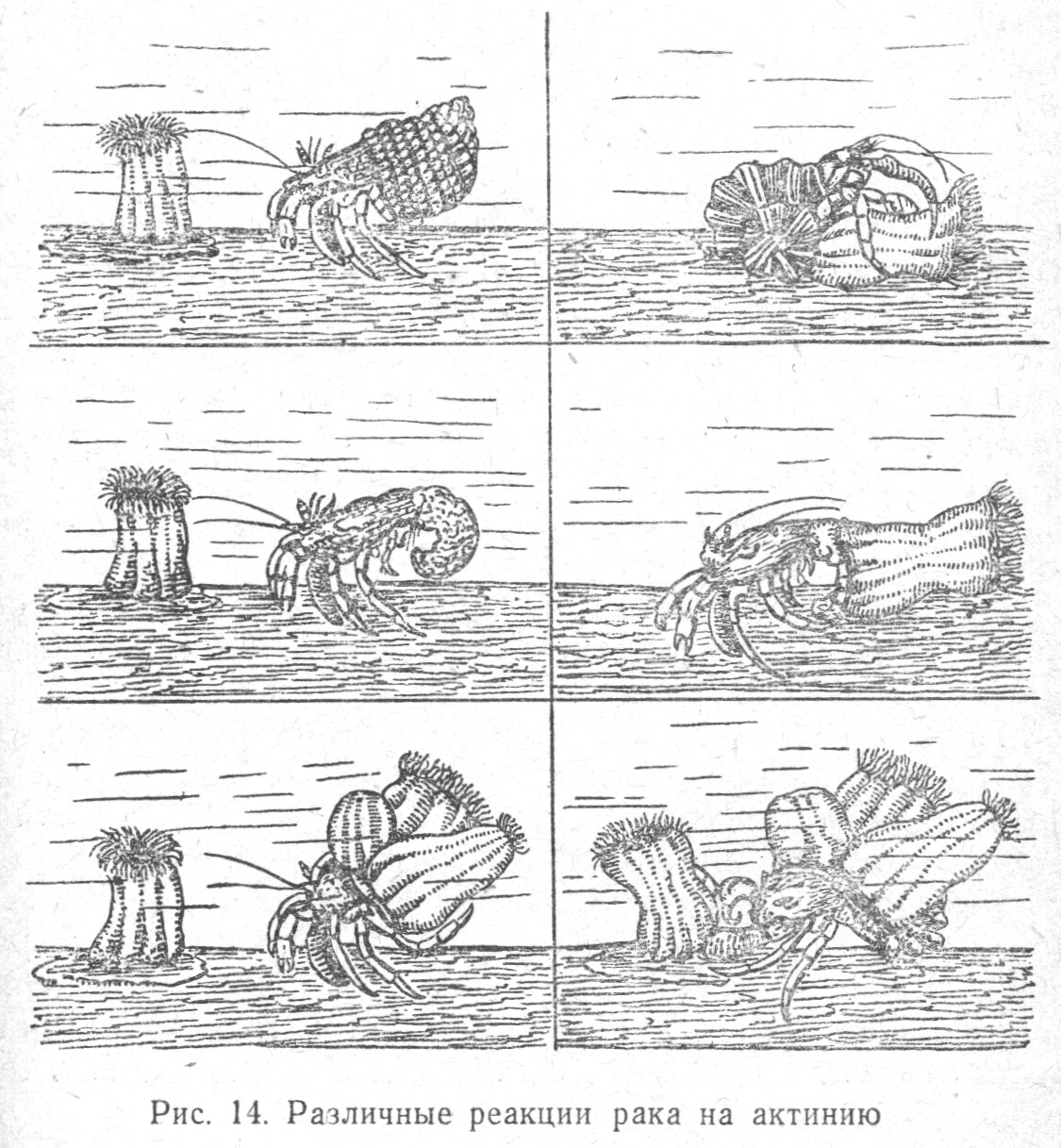
У ракообразных, в частности у рака-отшельника, инстинкт самозащиты выражен в том, что он использует раковины для помещения в них своего незащищенного панцырем мягкого брюшка. Вырастая, рак меняет раковины, предварительно ощупывая их полость усиками, клешнями и вынимая оттуда застрявшие твердые тела.

В дифференцировании ракообразными предметов разных форм, например, конических и плоских раковин, у них действует прежде всего осязательный, а не оптический раздражитель. Рак-отшельник, используя для защиты своего мягкого брюшка пустые раковины моллюсков, находя подходящую по величине тела раковину с отверстием, засовывает свое брюшко в отверстие, если оно расположено внизу раковины. Если же отверстие у раковины расположено вверху, то рак подрывает почву до тех пор, пока раковина не перевернется отверстием вниз, когда он может легко ее занять. При этом следует отметить, что рак-отшельник зрительно не различает раковины от круглого камня или деревянного шара, так как ощупывает и их, ища несуществующее отверстие. Для рака-отшельника каждый цилиндрический предмет имеет определенную биологическую значимость, но эта значимость различна в зависимости от состояния, в котором рак находится. Например, один и тот же вид животного (например, кишечнополостное — актиния) вызывает разные реакции рака (Табл. 1.13 и Табл. 1.14). Если брюшко рака защищено раковиной, — он берет актинию клешнями и насаживает ее на себя, тогда актиния, имеющая стрекатальные капсулы с обжигающей жидкостью, защищает рака от врагов. Если рака вынуть из раковины и поблизости не оказывается другой раковины — он пытается влезть внутрь актинии. Если рака долгое время не кормить, то он схватывает актинию и пожирает ее[50] (Табл. 1.14).
Таким образом, тот же предмет выступает для рака в различном биологическом значении, в зависимости от физиологического состояния животного: в первом случае — биологическая значимость актинии — «защитная», во втором — «жилищная», в третьем — «пищевая».
У членистоногих очень ярко выражен инстинкт ухода за молодью.
Самки крабов, например, прижимают ногами к брюшку отложенные ими яйца на время созревания икры.
Некоторые пауки (Lycosa) делают себе в земле глубокие, вертикально расположенные норки, вход в которые закрывают откидной крышечкой. Эта крышечка, слепленная из земли, глины и паутины, прикрепляется шарниром из паутины к стенке гнезда и легко откидывается при выходе паука из норки.
Водяной паук (Argyroneta aquatica) строит в воде колпак из пузырьков воздуха, принесенного с поверхности воды; под этот прозрачный колпак он и откладывает свои яички.
Самки некоторых видов наземных пауков делают для своих яиц кокон из паутины, который они носят под брюшком. После выведения паучков мать некоторое время носит их на своей спине, когда же паучата подрастают, каждый молодой паучок, отделяясь от матери, выпускает паутинку и остается таким образом связанным с ней; он возвращается к матери по этой же паутинке. Но по мере развития паучат связь между ними и матерью все более ослабевает и порой, особенно в неволе и при недостатке корма, паучата нападают на мать и, вонзая в нее челюсти, высасывают ее.
Анализируя взаимосвязи матери и детенышей, крупнейший знаток жизни и поведения паукообразных — профессор Вагнер[51] приходит к выводу, что у пауков обнаруживается периодически меняющаяся связь самки со своим потомством. У паучихи наблюдается как бы кривая развития и угасания инстинкта ухода за молодью в различные периоды развития молоди.
Сначала, когда яйца находятся в коконе, отношение самки к кокону нейтральное. Но ко времени выхода молоди из яичек сила инстинкта сохранения потомства доcтигает своего наивысшего развития. Например, если отнять кокон у самки тарантула (Theridium pictum) на седьмой или восьмой день после его изготовления, то паучиха с необычайной тщательностью начинает обыскивать свое помещение, перерывает всю землю и прекращает поиски лишь по возвращении ей кокона, который она схватывает и начинает носить. Более того, иногда паучиха подбирает искусственно подброшенные 2—3 кокона, носит их и охраняет до времени выхода молоди.
В первое время жизни паучат самка неотступно следит за ними, но чем более подрастает молодь, тем нейтральнее относится к ней мать, а позднее, ко времени полного созревания паучат, когда они становятся самостоятельными, она прерывает с ними всякую связь.
Таким образом, связь матери с потомством определяется биологической значимостью этой связи для обеспечения интересов вида на время беспомощности молоди, до возникновения у нее способности к самостоятельной жизни.
Наблюдая это весьма целесообразное поведение самок пауков, Вагнер определенно доказал ограниченность «материнского» инстинкта.
Нередко содержимое коконов уничтожают паразиты, тем не менее паучиха определенный срок (15—20 дней) носит опустевший кокон, а затем бросает его, даже не исследуя содержимого.
Иногда самка не откладывает яиц в кокон, а, посидев в той позе, в которой откладывает яйца, столько времени, сколько обычно требует откладывание яиц, тем не менее обращается к следующему этапу, т. е. заделывает пустой кокон, носит его с собой, как если бы он был наполнен яйцами.
Такова ограниченность и «слепота» инстинктивных действий, относящихся к уходу за молодью. Этот инстинкт, если проследить его у более примитивных до все более высокоорганизованных беспозвоночных животных имеет прогрессивное развитие, достигающее наивысшего уровня у колониальных насекомых.
В группе этих насекомых мы наблюдаем сложнейшие инстинкты, связанные с уходом за молодью и групповым поведением сочленов гнезда. Например, в колонии пчел имеется лишь одна плодущая самка, бо́льшая по размерам, так называемая «матка», она откладывает яйца, исчисляемые многими тысячами. Выходящих из неоплодотворенных яиц самцов- трутней пчелы после оплодотворения самок, к осени, убивают и изгоняют из улья.
Бесплодные рабочие-самки с недоразвитыми половыми органами — ухаживают за маткой, чистят улей, кормят молодь и собирают пищу.
Замечательно, что каждая рабочая пчела по мере своего развития последовательно меняет функции, начиная с более легких, связанных с кормлением личинок, и кончая более сложными операциями — полетами за пищей и постройкой ячеек.
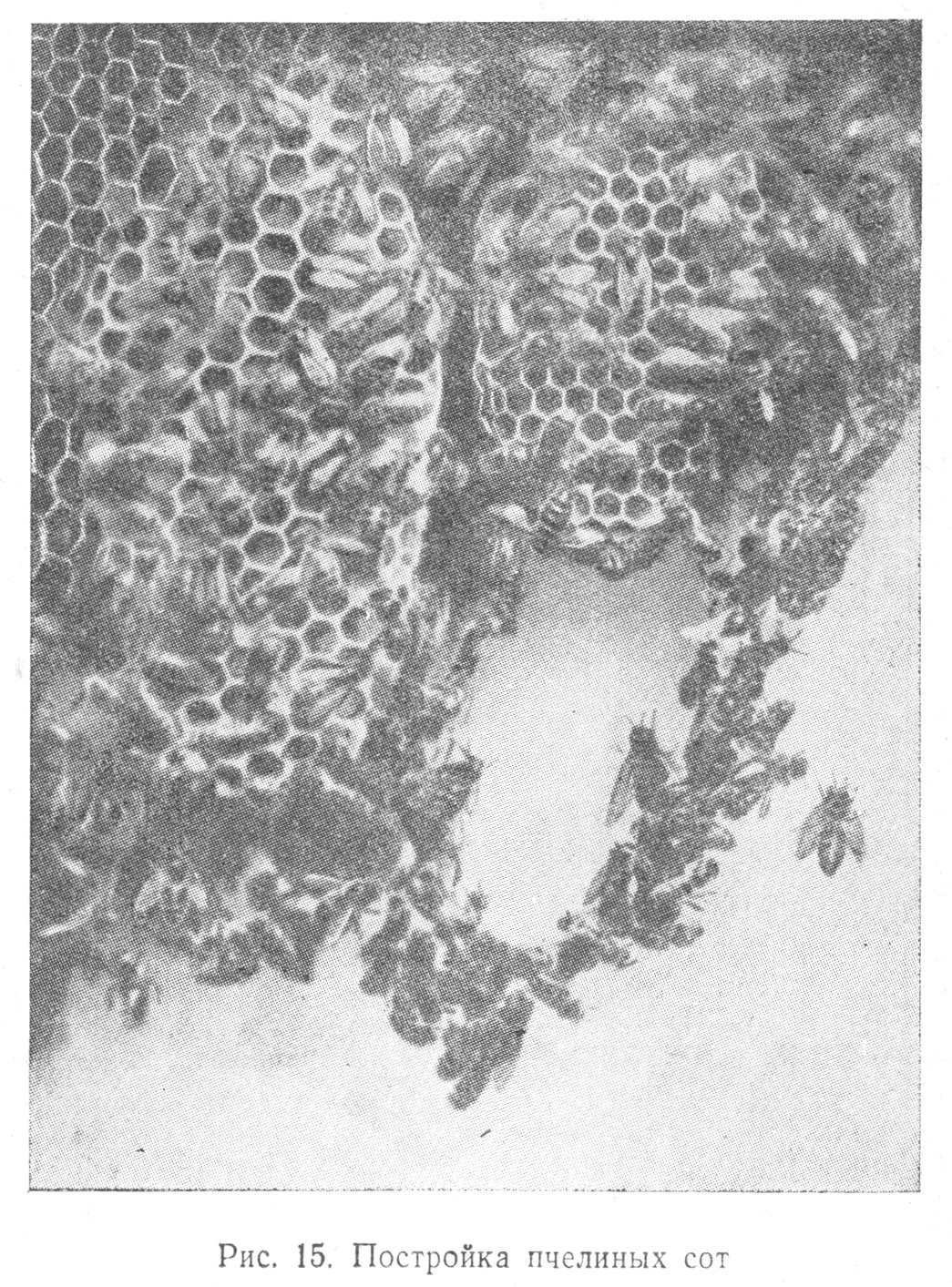
Всем известны изумительные постройки пчелиных сот с их сложными камерами, дифференцированными по расположению и по назначению: в одних складывается пища, в других помещаются яйца, личинки и т. д. (Табл. 1.15).
В случае опасности — при изменении температуры и влажности в камерах, где хранятся яйца, — последние переносятся в условия, более благоприятные для созревания молоди. Развивающиеся личинки пчел пользуются тщательным уходом рабочих пчел, соблюдающих определенный режим их питания.
При анализе проявления инстинкта ухода за потомством у представителей отряда перепончатокрылых насекомых мы можем явно наблюдать его эволюцию, начиная с той стадии, которая имеется у предков современных перепончатокрылых (сетчатокрылых), до образования первичной пчелиной семьи сфекоидных пчел, в которой я возникают сложные формы ухода за молодью, достигающие наивысшего уровня у настоящих пчел. Это изменение инстинктивных проявлений ухода за молодью, начиная от более простых до более сложных его форм, происходило под влиянием изменения условий жизни насекомых.
Углубленное исследование естественноисторического развития образа жизни и повадок этой группы насекомых энтомологом С. И. Малышевым вскрывает ряд последовательных фаз эволюции инстинкта ухода за молодью в отряде перепончатокрылых[52].
Согласно этому исследованию, оказывается, что предки современных перепончатокрылых (Hymenoptera), именно сетчатокрылые (Neúroptera), просто откладывали свои яйца в случайные места: в углубления, в щели, в почву, не проявляя о них никаких забот (у скорпионниц). Выходящие из яиц личинки вели бродячий образ жизни и самостоятельно добывали корм, питаясь близнаходящимися разлагающимися растениями, затаскивая их в вырытые норки. Следует отметить, что эти личинки малоподвижны и поэтому, очевидно, в процессе отбора могли выжить лишь те из них, которые получали со стороны родителей некоторый уход. Этот уход выражался в том, что матери начали прикреплять яйца к сочным зеленым частям растений, служащим пищей вылупившимся личинкам (у пилильщиков). Постепенно у материнской особи возникло еще большее усовершенствование приемов ухода за потомством: мать стала откладывать яйца под подрезанную ею кожицу листа, как в карман, тем самым предохраняя яйца от различных внешних вредоносных воздействий (также у некоторых видов пилильщиков).
На следующей стадии изменяются повадки личинок, вышедших из яиц, отложенных в поверхностный слой листовой ткани — личинки по вылуплении уходят в глубинные слои растений (листья, плоды, побеги), образуя всем известные вздутия или галлы (у пилильщиков). Теперь уже у особи-матери появляется инстинкт откладывания самих яиц в древесину хвойных и лиственных деревьев, в которых вылупившиеся личинки протачивают многочисленные ходы (у рогохвостов).
В связи с таким способом питания у личинок изменяется внешний вид: у них недоразвивается голова, грудные ножки и исчезают брюшные конечности.
Подобное изменение личинок вызывает появление новой формы материнского ухода за потомством. Этот уход проявляется в том, что особь-мать (из рода Isosoma inquilinae) изменяет растительный тип питания своих личинок на животный. Она начинает откладывать яйца не на любые стебли растений, а на поврежденные галлами растения, заключающие в себе личинки наездников другого вида (Isosoma rossicum).
Выходящая личинка — Isosoma inquilinae — переходит на смешанное питание, пожирая и выходящую из галла личинку, и стенки галла, и ткани растения.
Возникает явление паразитизма одного вида насекомых за счет другого и плотоядное питание личинок.
В следующей стадии ухода за потомством у самок наездников наблюдается повадка откладывать яйца не близ галла, а на выходящую из него личинку не только близкого к себе вида, но и личинок других насекомых (жуков-златок), даже на яйца насекомого хозяина (например, одиночных пчел — Osmia parvula). Вледствие этого выходящая из яйца личинка-гостья пожирает не только яйцо хозяина, но и заготовленное ему медвяное тесто.
Еще позднее наступает так называемая эктопаразитическая стадия в развитии инстинкта ухода за молодью, т. е. самки начинают открыто нападать на свои жертвы, независимо от места их обитания (на растениях или на поверхности земли), раня их и откладывая яйца под кожно-мускульные покровы тела жертвы.
Наконец, мать парализует живую, энергично сопротивляющуюся добычу, пронзая ее яйцекладом, получившим функцию жала.
Дальнейшее осложнение материнского инстинкта ухода за потомством выражалось в прятании жертвы, на которой самка откладывала свои яйца, в защищенные места (в древесину растений, в землю, в норку), что было связано с вырыванием норки, с закрыванием ее крышкой, предохраняющей от доступа к ней насекомых-врагов и от различных внешних вредоносных воздействий; это явление наблюдается у ос. На этой стадии уход за потомством уже включает 5 актов:
нахождение живой добычи и ее парализация;
устройство в земле норки для помещения добычи;
перенос добычи в норку (Табл. 1.16);
откладывание яйца на добычу;
зарывание норки с помещенной в ней добычей и яйцом.
На следующем этапе развития инстинкта ухода за потомством обнаруживаются попытки увеличить запас приносимой матерью для личинки пищи: не одной, а двух жертв, хотя и меньшего размера; оса начинает откладывать яйца в норку до помещения добычи, постепенно создавая запас по мере роста личинки (сначала из одной жертвы, потом двух); самка питает при этом личинку разжеванной добычей.
На новом этапе развития инстинкта ухода за потомством появилось построение матерью самостоятельной ячейки с применением строительного материала (осиной бумаги). Так возникла постройка сота и стало возможным откладывание в соты тысяч яиц, хорошо защищенных извне, выкармливание осой своих личинок плотоядной пищей, разжеванной самой матерью (ново-осиная стадия). Еще позднее мать-кормилица перешла от плотоядного к растительному корму, питаясь нектаром и пыльцой цветов.

У некоторых перепончатокрылых имеются и осиные, и пчелиные повадки в выкармливании молоди. На этой базе возникли и собственно пчелиные повадки в развитии: инстинкта ухода за потомством.
Теперь яйца сначала временно откладываются в глубине выводковой камеры, а после заготовки медвяного теста помещаются на него; далее медвяная кашка прикрепляется к стенкам ячейки (у наиболее примитивных коллетовых пчел — Colletidae); еще далее (у сфекоидных пчел)[53] забота о потомстве выражается в следующих 5 актах:
устройство ячейки;
заготовка корма;
откладывание яйца на стенки ячейки (а не на корм);
складывание в ячейку корма, выделенного изо рта;
выделение изо рта на стенки ячейки жидкости — секрета, защищающего корм от вредоносного воздействия.
В последующем этот инстинкт значительно осложняется. Теперь оказывается, что изоляционная пленка в ячейке настилается ранее складывания медвяного корма, а потом яйцо прикрепляется к потолку ячейки, так как в глубине ячейки для него не остается места. Так образуется первичная полупчелиная (сфекоидного типа) фаза развития сложного инстинкта ухода за потомством у перепончатокрылых.
Обзор эволюции этого инстинкта совершенно явно обнаруживает, что от одноактной стадии ухода за потомством идет ряд постепенных осложнений к 2—6 и еще более многоактной стадии.
Одноактная стадия. Самка откладывает яйца в углубления, щели, ямки на поверхности растений (у сетчатокрылых — скорпионниц).
Двухактная стадия. Самка подрезает кожу листьев, просверливает древесину, или кожно-мускульный покров жертвы, и откладывает яйца внутрь (у пилильщиков, у рогохвостов, у Isosoma inquilina).
Трехактная стадия. Парализация живой добычи, пробуравливание ее покрова, откладывание яиц под покров жертвы (у ос).
Четырехактная стадия. Парализация живой добычи, унос и прятание ее, пробуравливание ее покровов, откладывание яйца (у наездников).
Пятиактная стадия. Парализация живой добычи, выкапывание норки, перенос жертвы в нору, откладывание яиц на жертву, закрывание норы (у ос).
Семиактная стадия. Выкапывание норки, закрывание норки временной крышкой, нахождение и парализация жертвы и откладывание на ней яйца, открывание временной крышки, вмещение в норку жертвы с отложенным на ней яйцом, окончательное закрывание норки (у ос).
Десятиактная стадия. К шести вышеперечисленным актам присоединяются три новых: двукратное закрывание гнезда, двукратная охота и двукратный перенос жертвы (у ос).
Многоактная стадия. Возникает замедленный последовательный многократный принос матерью разжеванной ею самой пищи для личинки (у ос).
Далее наступает более сложная многоактная стадия: возникает постройка из осиной бумаги гнезда, откладывание тысячи яиц, при последовательном снабжении пищей, разжеванной матерью, тысяч личинок (у ос).
Происходит не только количественное нарастание многоактности цепных инстинктивных действий, но осуществляются и качественные изменения форм ухода за потомством, увеличивающие его эффективность. Эти качественные изменения, включающие более сложные формы отношений матери с потомством, связаны с развитием более сложных форм отражения внешних раздражителей. Если в начальных одноактных стадиях, ухода за потомством мать ограничивается лишь восприятием места, материала, живой добычи для помещения на нее своего яйца, то в последующем она не только находит подходящий субстрат для помещения яиц, но несколько подрабатывает его, надрезая, полуразрушая кожицу, под которую кладет яйцо (двухактная стадия).
Далее происходит выделение, связанное с различением особью-матерью подходящего субстрата от неподходящего, что требует более точного сенсорного дифференцирования — выбора не любого стебля растения, а стебля с наличием галла.
Далее у матери возникает способность к отыскиванию глубоко запрятанного живого организма для помещения на него яйца, что требует уточнения сенсорных зрительных и обонятельных дифференцировок. Еще далее у самки расширяется сфера поиска жертв, а в связи с этим — ориентировочно-исследовательский рефлекс и зрительная дифференцировка мест обитания добычи, что сочетается с преимущественным, весьма адаптивным использованием скрыто живущей добычи, т. е. личинок (трехактная стадия).
В недрах прежнего комплекса приемов ухода за потомством у особи-матери возникает качественно новый активный прием, связанный с овладеванием жертвой, с парализацией живой добычи, что опять-таки сопровождается появлением ряда новых количественных и качественных изменений инстинкта (т. е. 4-5-7-10-актных стадий). В процессе ухода за потомством участвует целый комплекс рецепторов (обонятельных, зрительных, осязательных) и эффекторов. Несомненно, что мы наблюдаем поступательный характер развития инстинкта ухода за потомством; возникновение новых сложных связей, комбинаций многих рефлексов в единую сложную цепь.
Совершенствуется, становясь все более и более активным, прием сохранения и прятания парализованной жертвы, включающий уже не один, два, три четыре акта, а осуществление четырех, пяти, семи-десяти актов.
Это прятание жертв связано не только с роющей, разрушительной деятельностью насекомого, но и с конструктивной — оформление одной крышки норки, а позднее двух крышек; затем насекомое включает новый строительный материал (осиную бумагу) при наличии многоактной стадии.
Практический анализ и практический синтез при изменении деятельности насекомого все более усложняется и уточняется. Аналитико-синтетическая деятельность насекомого выражается в избирательном последовательном выделении определенных раздражителей, имеющих сигнальное значение, и в их синтезировании в сложные комбинации, цепи безусловнорефлекторных связей.
Возникновение качественно новой реакции — акта постройки осиного гнезда, создает возможности для новых количественных изменений, т. е. откладывания многочисленных яиц, выведения многочисленного потомства, соединенного в общество (ново-осиная фаза).
Но наличие огромного количества потомства, необходимость его выкармливания, естественно, требовала от особи-матери чрезвычайно большой активности, связанной с нахождением живой добычи, ее умерщвлением, борьбой с ней.
Необходимость выкармливания многочисленной молоди и затруднительность выкармливания ее живой добычей приводит к возникновению иного, не плотоядного кормления личинки, а более легкого для осуществления растительного кормления. Это последнее дает возможность увеличить количество заготовляемой матерью пищи настолько, что она создает склад медвяной кашицы и выстилает пленкой выводковые камеры для помещения в них яиц.
Но здесь наступает новое осложнение: при обилии провизии в глубине, внутри ячейки не остается места для развития личинок. Это затруднение разрешается тем, что мать начинает откладывать яйцо на потолок ячейки и не ранее, а после заготовки личиночного корма (первично-пчелиная фаза сфекоидного типа).
Далее возникают те изумительные формы ухода за потомством, которые мы наблюдаем у колониально живущих настоящих пчел.
Таким образом, можно проследить эволюцию сложного инстинкта ухода за потомством у перепончатокрылых, достигающего высокого уровня у настоящих пчел, у которых имеются уже отмеченные нами замечательные формы ухода рабочих пчел за яйцами, за вылупляющимися личинками, выкармливание личинок питательными веществами, вырабатываемыми особыми железками, молочком, пыльцой и нектаром, собранными пчелами с цветов, запечатание ячеек с личинками, закончившими свое развитие.
Таковы данные, с полной определенностью убеждающие нас в наличии эволюции инстинктов в группе перепончатокрылых насекомых, данные, наглядно показывающие нам, как более простые безусловнорефлекторные акты, относящиеся к выведению молоди, в связи с изменением образа жизни, способа питания и приемов обеспечения пищей личинок насекомых приводили к образованию все более и более сложных комбинаций инстинктивных действий.
Длинные цепные системы последовательно и преемственно смыкающихся актов образовали сложные формы инстинктивного поведения, связанного с уходом за потомством. В этой эволюции совершенно очевидным оказывается расширение круга деятельности насекомых, развитие и совершенствование форм отражения, совершенствование аналитико-синтетических процессов в филогенезе организмов.
Без знания постепенно слагающихся форм поведения существование этих замечательных инстинктов в их готовом виде казалось бы чудом творчества природы, и только учет их постепенного возникновения при ведущем воздействии внешней среды на организм, в силу единства организма с условиями его существования, дает разгадку этого чуда. Это объяснение сложнейших форм проявления инстинктивной деятельности животных, в противовес давним теологическим и телеологическим интерпретациям проблемы инстинкта, как и различным новым неправильным ее освещениям в зарубежной литературе, базируется на материалистическом его обосновании советскими учеными.
И. П. Павлов весьма осторожно указывал на возможный путь происхождения инстинктов: безусловные рефлексы возникают из условных, повторяющихся из поколения в поколение. Он писал: «В высшей степени вероятно (и на это имеются уже отдельные фактические указания), что новые возникающие рефлексы, при сохранности одних и тех же условий жизни в ряде последовательных поколений, непрерывно переходят в постоянные» и далее: «можно принимать, что некоторые из условных, вновь образованных рефлексов позднее наследственностью превращаются в безусловные»[54].
Возникнув под воздействием внешней среды и будучи явлением приспособительным к строго определенным условиям, инстинктивное поведение в известной мере стереотипно и при резком отклонении условий от нормальных обнаруживает свою несостоятельность. Замечательные опыты выдающегося наблюдателя и знатока жизни насекомых Фабра подтверждают это положение многочисленными фактами.
Так, например, пчелы заполняют медом и нарочито продырявленную человеком ячейку; осы замуровывают осмотренные ими пустые норки, хотя человек перед тем извлек оттуда положенную осой гусеницу (предназначенную для корма будущей личинки) с отложенным на ней яйцом осы.
Обычно, вырыв норку, земляная оса закрывает ход в нее камешком, как бы маскируя его; прилетев с добычей, она кладет ее у входа, отодвигает камешек и еще раз контролирует норку. Если же за то время, пока оса находится в норке, ее добычу отодвинуть, то, поискав и найдя парализованное насекомое, она все-таки опять оставляет его у норы и предварительно опять обследует нору, хотя в этом уже нет надобности.
Фабр 40 раз отодвигал добычу (сверчка) от норки осы и тем не менее оса каждый раз обыскивала норку, прежде чем стащить в нее жертву[55] . Такова ограниченность инстинкта.
Сложная общественная жизнь насекомых вызвала появление своеобразных способов их взаимного общения, выражающихся в движении усиков (у пчел и муравьев), в изменении силы и ритма их ударения, дрожании головы (у термитов), «танцах» пчел[56].
Наблюдения и опыты Фриша показали, что пчелы-сборщицы, возвращающиеся с мест кормления, «сообщают» пчелам, находящимся в улье, о расстоянии, на котором они обнаружили богатый источник корма.
Если пчелы вернулись с близкого расстояния, то они производят круговой танец, т. е. кружатся на одном месте, сначала направо, а затем налево, неоднократно повторяя эти круговые движения в течение полминуты и дольше. Другие пчелы, видя «танцы», приходят в сильное возбуждение, толпой следуют за «танцующей» пчелой, касаясь своими антеннами ее тела. Далее эти пчелы внезапно вылетают из улья, направляясь к источнику корма, а по возвращении в улей также начинают «танцевать», сигнализируя о наличии корма. Обнаружив источник корма, находящийся на далеком расстоянии от улья (например, 300 метров), пчелы исполняют «виляющий танец». В этом случае после небольшого пробега по прямой линии, они начинают быстро вилять брюшком из стороны в сторону; затем они делают поворот налево, на 360°, потом снова бегут по прямой и делают полный поворот направо. Подобного рода движения повторяются многократно. Фриш экспериментально доказал, что «виляющий танец» сигнализирует не только о дальности расстояния от улья источника корма, но и о том, насколько велико это расстояние. Это последнее определяется числом поворотов пчелы, выполняющей «танец». Фриш подчеркивает, что по разным формам танца обитатели улья явно узнают, на какое расстояние им надо лететь, чтобы добраться до корма.
Кроме того, при более детальном анализе этих явлений Фриш показал зависимость направления прямолинейной части виляющего танца от положения солнца.
Если прямолинейная часть виляющего танца направлена прямо вверх — это означает, что источник корма находится в том же направлении, что и солнце, если же прямолинейная часть виляющего танца направлена вниз — то это значит, что корм находится в противоположной стороне от солнца. Более того, если прямолинейная часть этого танца направлена под углом в 60° от направления силы тяжести — то это значит, что место кормежки пчел расположено влево от солнца, если же на 120° вправо — то это значит, что источник корма находится на 120° вправо[57].
Кроме того, у пчел имеется и звуковая сигнализация в виде жужжания, дребезжания, издавания подобия трубных звуков, вызывающих определенные реакции сочленов группы. Конечно, эти движения и звуки — лишь внешнее выражение тех или других состояний возбуждения насекомых, но никоим образом не предметные знаки общения: тем не менее, объективно они являются определенными (хотя и не направленными) сигналами для сочленов своего вида, и вызывают соответственные их реакции.
Для членистоногих характерна не только богато развитая инстинктивная деятельность, но и способность использовать индивидуально приобретенные опыты на основе образования условнорефлекторных связей.
Проводились исследования способности краба к ориентировке в лабиринте, одна часть которого вела к выходу, другая — к тупику; эти опыты обнаружили, что если в первых 10 опытах правильная ориентировка краба составляла лишь 50%, то в последующем обнаружилось улучшение результатов нахождения правильного выхода, повышение процента положительных решений до 60, 75, 83, 90 (Табл. 1.17)[58].
После двухнедельного перерыва в работе количество правильных решений крабами той же задачи уменьшилось. Обученные в лабиринтах омары, делавшие после тренировки одну ошибку из 10 опытов, после перерыва в занятиях с ними в тех же 10 опытах делали уже три ошибки; это означало, что хотя условный рефлекс у омаров и сохранялся, но он оказывался весьма непрочным. Была доказана способность краба и к переучиванию при нахождении свободного пути в лабиринте, идущего вместо правой, по левой его стороне, причем было установлено, что, отыскивая путь, крабы пользовались зрением, осязанием и мышечным чувством.
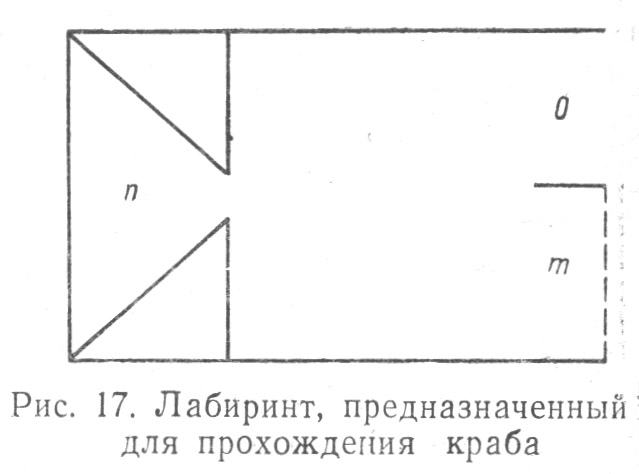
При постановке сравнительных опытов ориентировки в лабиринте краба и других животных обнаружилось, что ориентировочная способность краба при нахождении кратчайшего пути к выходу значительно ниже, чем таковая муравьев и мыши.
У раков-отшельников удалось образовать ассоциативную связь двух разных раздражителей: положительного — химического и отрицательного — светового, располагая в тени излюбленную раками пищу.
В первый день из 30 раков через 15 минут пищу нашли 3 рака, в третий — 20 раков, на восьмой день из 29 раков через 15 минут нашли пищу 28 раков; на тринадцатый — из 27 раков через 5 минут нашли пищу 25.
Таким образом, мы замечаем, как у раков постепенно вырабатывался условный рефлекс на нахождение пищи, и не имевший ранее отношения к пище раздражитель — тень в результате сочетания с кормлением стал положительным пищевым сигналом.
У насекомых отмечена хорошая память на места и способность возвращаться к дому.
Обычно, отлетая от гнезд, молодые пчелы и осы делают вокруг гнезда круговые полеты; эти полеты, все больше и больше расширяющиеся в диаметре, помогают пчелам зрительно запоминать вехи, находящиеся близ гнезд, и когда гнездо перемещают, то пчелы при перелете либо кружатся на месте бывшего гнезда (при отсутствии прежних вех), не находя свое гнездо, если же вехи, например, большие камни, снова кладут близ гнезда, пчелы немедленно находят гнездо. Как мы уже упоминали, у насекомых отчетливое различение формы предмета возможно лишь на близком расстоянии; далеко расположенные предметы имеют для них весьма расплывчатое очертание.
Превосходно доказана способность насекомых под влиянием изменившихся условий, например, воздействий со стороны человека, изменять свое прирожденное, инстинктивное поведение, подчиняться дрессировке и вырабатывать довольно сложные навыки, основанные на образовании условных рефлексов.
Так, например, таракан, обычно отрицательно фототропичное насекомое, был приучен оставаться на свету, после того как его пребывание в темноте связали с электрическими ударами, шоком. Но это выработанное поведение сохранялось только 55 минут, по истечении которых таракан снова шел в тень; однако, новая выучка при воздействия тех же стимулов наступала значительно быстрее.
Ос выдрессировали подходить к экспериментатору, который поглаживал их; жук- плавунец выучился взбираться за куском мяса по палке, опущенной в воду в определенном месте; муравьи легко ориентировались в нахождении правильного пути в сложном лабиринте, имевшем, кроме свободного выхода, несколько (5—10) тупиков; пчел удалось приучить прилетать к ящику определенного (например, голубого) цвета, или выбирать коробку с изображением над ее летным отверстием определенной формы цветка (см. Табл. 1.10).
Дрессировка насекомых (пчел) имеет большое сельскохозяйственное значение.
В практических целях пчел приучили посещать цветы красного клевера, обычно мало посещаемые ими, что повысило сбор меда и содействовало опылению этой лучшей из кормовых трав, обеспечивающей, кроме того, обогащение азотом почвы, на которой клевер произрастает[59].
В целях повышения перекрестности опыления удалось искусственным путем переключать пчел с посещения растения сильного медоноса, например, гречихи, на растение, бедное нектаром и слабо посещаемое пчелами (горчицу)[60].
В этих случаях приходилось применять в ульях двустороннюю выработку условных рефлексов; угашать двигательную реакцию прилета насекомого к сильному медоносу путем применения в улье отвергаемого вещества (раствора лимонной кислоты), настоенного на цветах сильного медоноса, обладающего запахом. Кроме того, в улье производилась подкормка пчел сиропом, настоенном на слабом медоносе, также обладающем своим специфическим запахом. Таким образом удалось снизить посещаемость пчелами сильного медоноса и переключать их на посещение слабого медоноса.
Эти опыты показали, что, комбинируя соответствующим образом положительное пищевое подкрепление и отвергаемое вещество в сочетании с сигнальными признаками того или другого растения, можно управлять поведением пчел, усиливая или затормаживая их прилеты на те или другие цветы.
В итоге обзора и анализа рецепции и поведения членистоногих, мы можем сделать определенный вывод о повышении уровня отражения этими животным окружающего мира, по сравнению с наблюдаемым у кишечнополостных и червей. Членистоногие многообразнее, дробнее, точнее и тоньше отражают окружающее.
Рецепция членистоногих обогащается, во-первых, за счет дифференцировки ранее обобщенной химической рецепции, совмещающей обонятельную и вкусовую, на раздельно представленную: обонятельную и вкусовую.
Зрительная рецепция дает возможность различения цвета, формы, движения предметов, в то время как представители животных более низкоорганизованного типа (черви) различают лишь свет, тень и направление света. У насекомых возникает предметное, хотя и ситуационно связанное восприятие.
Впервые в филогенезе организмов у членистоногих появляется звуковосприятие — фонорецепция и способность к произведению звуков.
Все эти особенности рецепторики, конечно, указывают на то, что аналитико- синтетическая деятельность членистоногих; в частности насекомых, поднимается на более высокий уровень, чем то имело место у вышерассмотренных типов животных (кишечнополостных и червей).
Осуществление анализа новых видов энергии, связанного с возникновением новых видов рецепции и появлением новых анализаторов, трансформирующих внешнюю энергию в нервный процесс, предполагает увеличение анализаторной способности членистоногих.
У членистоногих возникают реакции не только на элементарные раздражители, отдельные признаки предметов, но и на комплексы раздражителей, на сложные образы.
Появление у членистоногих зачатков сложных целостных восприятий свидетельствует о наличии слияния отдельных раздражений в единый комплекс предмета, указывая на высокое развитие аналитико-синтетической деятельности их мозга. Напомним о замечательной конструктивной деятельности паукообразных и особенно насекомых — сооружение тенет, ловушек, гнезд, коконов, — об употреблении предметов в опосредствованном значении у ракообразных — самомаскировки крабов с привлечением предметов окружающей среды — водорослей, губок, актиний.
Тончайшая аналитико-синтетическая деятельность проявляется в дифференцировании колониальными насекомыми биологически значимых (полезных и вредных) раздражителей и в соответственных высоко приспособленных многообразных реакциях на них. Напомним различение этими насекомыми сочленов своих гнезд от чужаков, приемы ухода за яйцами, в зависимости от изменения температуры и влажности в улье и т. д. и т. п.
В обзоре эволюции поведения беспозвоночных животных мы отметили не только количественное его усложнение, но появление и качественно новых форм, коренных изменений в старых, исторически сложившихся инстинктах.
Движущей силой этих изменений оказывалось возникновение противоречивого соотношения между организмом и условиями его существования, приводящего к борьбе этих противоречий, разрешающихся в процессе приспособительного уравновешивания организма с условиями его существования.
Хотя практический синтез у членистоногих, в частности осуществляемый в сложных формах строительной конструктивной деятельности (например, при постройке тенет, ловушек, гнезд), свидетельствует о довольно высокой интегрирующей функции головных ганглиев, все эти: действия являются инстинктивными и принципиально отличаются от осмысленных, сознательных действий. Они: представляют сложные цепи безусловнорефлекторных актов, в которых запечатлелся видовой (т. е. филогенетический) опыт животного, осуществляемый при узко ограниченных, строго определенных условиях окружающей среды.
Как справедливо отмечает Маркс, «паук совершает операции, напоминающие операции ткача, и пчела постройкой своих восковых ячеек посрамляет некоторых людей — архитекторов. Но и самый плохой архитектор от наилучшей пчелы с самого начала отличается тем, что прежде чем строить ячейку из воска, он уже построил ее в своей голове. В конце процесса труда получается результат, который уже в начале этого процесса имелся в представлении работника, т. е. идеально»[61].
Это положение как нельзя лучше документируется тем, что при некотором изменении условий гнездостроения пчел, например, при нарочитом продырявливании пчелиной ячейки, пчелы не заделывают ее, не замечая дефекта и наполняют ее медом, как если бы она была целой.
Строя ячейки, они реагируют только на безусловные сигнальные, нормально действующие раздражители, так как их восприятие узко ситуационно связано, и их эффекторные реакции на определенные раздражители воспроизводятся в той связи и последовательности, как это требуется согласно сложившемуся стереотипу данной формы инстинкта. Сразу сломать рамки стереотипа, преодолеть косность инстинктивных действий, затормозить воспроизведение нецелесообразных действий, заново перестроить инстинктивное поведение в соответствии с появлением новых раздражителей насекомому трудно. Однако, насекомое может преодолевать ограниченность инстинкта выработкой условных рефлексов.
Наблюдения в природе и лабораторные опыты убедительно доказывают наличие у членистоногих анализа и синтеза раздражителей, основанного и на использовании ими своего индивидуального опыта.
Напомним выработку условных рефлексов путем установления связи между безусловным химическим раздражителем (пищей) и условным световым раздражителем (тенью) у ракообразных.
Для насекомых условными сигнальными раздражителями, которые животное приучилось дифференцировать от других, были запах, цвет, форма и др. Эти новые условные раздражители выделялись насекомыми из комплекса раздражителей, одновременно с ними присутствующих в окружающей обстановке. Эти раздражители вступили в связь и синтезировались с безусловными (пищей).
Образованные условные связи членистоногих отличались большой прочностью и могли сохраняться после перерыва в их воспроизведении. Это указывало на сохранение следов от раздражения, на наличие памяти у членистоногих, на известную пластичность их инстинктов, на подвижность форм отражения ими среды.
Эта возможность значительного обогащения «рефлекторного фонда насекомых» в связи с меняющейся жизненной обстановкой дает право на признание более высокого развития психики у этих форм беспозвоночных животных по сравнению с ранее рассмотренными.
[18] В. А. Павлов. Раздражимость и формы ее проявления. «Советская наука» 1954, cтр. 105.
[19] А. А. Заварзин. Очерки по эволюционной гистологии нервной системы. Избр. труды, т. III, изд. АН СССР, 1950, стр. 18 и далее.
[20] А. А. Заварзин. Избр. труды, т. III, изд. АН СССР 1950, стр. 24—25.
[21] Там же, стр. 27—28.
[22] И. П. Павлов. Полн. собр. соч., т. III, кн. 1, изд. АН СССР. М, 1951, стр. 124—125.
[23] Л. Г. Воронин. Анализ и синтез сложных раздражителей у высших животных. Медгиз, 1952, стр. 26.
[24] И. П. Павлов. Полн. собр. соч., т. III, кн. 2, изд. АН СССР, М., 1951, стр. 24.
[25] В. А. Павлов. Раздражимость и формы ее проявления. «Советская наука», 1954, стр. 107.
[26] А. А. 3убков и Г. Г. Поликарпов. Условный рефлекс у кишечнополостного животного. Журн. «Успехи современной биологии», 1951, т. XXXII, вып. 2 (5), стр. 301—302.
[27] Ч. Дарвин. Образование растительного слоя деятельностью дождевых червей. Изд. Поповой, С.-П., 1899 (пер. с англ.).
[28] О. Mangold. Beobachtungen und Experimente zur Biologie d. Regenwurm. Z. f. vergleichende Physiologie. Heft 2. 1924.
[29] В. А. Вагнер. Сегментарная психология. Серия «Новые идеи в биологии» Сб. 6-й биопсихология, изд. «Образование», С.- П., 1914, стр. 38—138.
[30] R. М. Yerkes. The intelligence of earthworms, J. of Animal Behavior, v. 2, № 5, 1912.
[31] См. В. М. Боровский. Психическая деятельность животных. Биомедгиз, 1936.
[32] F. Hempelmann. Tierpsychologie. Akad. Verlagsgesellschaft. Lpz, 1926, S. 222.
[33] К. Фриш. Пчелы, их зрение обоняние, вкус и язык. ИЛ, М., 1955, стр. 27.
[34] К. Фриш. Пчелы, их зрение, обоняние, вкус и язык. ИЛ., М., 1955, стр. 22—23.
[35] Джон Леббок. Муравьи, пчелы, осы. Перевод с англ., М., 1898, стр. 284.
[36] К. Frisсh. Der Farbensinn und Formensinn der Biene, Jena,1914.
[37] К. Фриш. Пчелы, их зрение, обоняние, вкус и язык. ИЛ, М., 1955, стр. 22—23.
[38] J. Uexküll. Streifzüge durch die Umwelten von Tieren und Menschen. В., 1934.
[39] В. А. Вагнер. Биологические основания сравнительной психологии, т. I, изд. Вольф, 1910, стр. 217.
[40] И. П. Павлов. Полн. собр. соч., т. IV, изд. АН СССР. М., 1951. стр. 23—24 (курсив наш — Н. Л.-К.).
[41] Там же, стр. 25.
[42] Там же, стр. 23.
[43] Там же, стр. 26.
[44] К. М. Быков и А. Т. Пшоник. О природе условного рефлекса. «Физиологический журнал СССР», № 5, XXXV, 1949, стр. 510.
[45] Е. В. Пузанова-Малышева. Муравьиные львы и их ловчие воронки. Сб. «Труды института эволюционной физиологии и патологии высшей нервной деятельности им. И. П. Павлова», т. I, изд. АН СССР, 1947, стр. 271.
[46] Fr. Doflein. Der Ameisenlöwe. Jena, 1916. S. 138.
[47] E. В. Пузанова-Малышева. Муравьиные львы и их ловчие воронки. Сб. «Труды института эволюционной физиологии и патологии высшей нервной деятельности им. И. П. Павлова», т. I, изд. АН СССР, 1947, стр. 282.
[48] О наличии изменений инстинктов и о типах этих изменений весьма обстоятельно писал еще В. А. Вагнер в своей классической монографии «Биологические основания сравнительной психологии», т. I, изд. Вольф, С.-П., 1910, стр. 55—93.
[49] R. Minkiеwiсz. Versuch einer Analise des Instinkts, Zool Jahrb. Abt. f. Syst., Bd. 28, 1910.
[50] J. Uexküll. Streifzüge durch die Umwelten von Tieren und Menschen. В., 1934.
[51] В. А. Вагнер. Возникновение и развитие психических способностей, вып. 9. Психология размножения и ее эволюция. Изд. «Начатки знаний», Л., 1929, стр. 59—64.
[52] С. И. Малышев. Пути и условия эволюции инстинктов низших перепончатокрылых (Symphyta и Terebratia): «Журнал общей биологии», т. X, № 1, изд. АН СССР, 1949, стр. 1—41.
С. И. Малышев. Пути и условия эволюции осообразных перепопчатокрылых (Vespoidea и Sphecoidea). ДАН СССР т. LXV, 1949, № 4, стр. 507—570.
С. И. Малышев. Пути и условия происхождения пчелиных (Hymenoptera Apoidea). ДАН СССР, т: LXXII, 1950, № 5, стр. 969—972.
[53] С. И. Малышев. Пути и условия происхождения пчелиных (Hytnenoptera Apoidea), ДАН СССР. Новая серия, т. LXXII, 1950, № 5, стр. 971.
[54] И. П. Павлов. Избр., труды. Изд. АПН РСФСР, М., 1951, стр. 215; 217. Однако см. также письмо И. П. Павлова от 1 марта 1927 г., опубликованное в газете «Правда» № 106 (3638), от 13 мая 1927 г. Раздел «Библиография», стр. 5.
[55] К. Фабр. Инстинкты и нравы насекомых. Изд. А. Ф. Маркса, С.-П., 1906, стр. 45—46 (пер. с франц.).
[56] К. Фриш. Пчелы, их зрение, обоняние, вкус и язык. М. И. Л., 1955, стр. 83.
[57] К. Фриш. Пчелы, их зрение, обоняние, вкус и язык. М. И. Л., 1955, стр. 63.
[58] R. М. Yerkеs, G. Е. Guggins. Habit formation in the craufish, Cambarus affinis. Harward Psych. Studies, v. I, 1903.
[59] Опыты проф. А. Ф. Губина. Медоносные пчелы и опыление красного клевера. ОГИЗ. Сельхозгиз, 1947.
[60] А. К. Воскресенская и Н. Г. Лопатина. Пищевые и оборонительные условные рефлексы у пчел. Труды института физиологии им. И. П. Павлова, т. II, 1953, стр. 542—561.
[61] К. Маркс. Капитал, т. 1. Госполитиздат, 1955, стр. 185.