Класс амфибий представляет первый этап на пути перехода позвоночных от водного существования к наземному. Только в личиночном состоянии и во время размножения амфибии живут в воде и дышат жабрами, взрослые особи дышат легкими и живут на земле, близ воды (бесхвостые амфибии) или преимущественно в воде (хвостатые амфибии — тритоны и др.).
Бесхвостые амфибии (лягушки, жабы) днем держатся обычно в затененных местах, под камнями или в зарослях прибрежной растительности, выходя на свет в сумерки для ловли живой добычи.
Изменение водного образа жизни и переход к полуводному и полуназемному, как то имеет место у амфибий, привело к изменению внешнего, вида, строения тела, мозга, органов чувств и основных типов поведения этих животных.
Тело амфибий более расчленено, чем тело рыбы, — амфибии уже имеют голову, обособленную от туловища, сильно выступающие по бокам тела подвижно соединенные с ним передние и задние конечности (ноги) с расчаленными пятью пальцами.
Хотя части головного мозга у амфибий, как и у рыб, вытянуты в один ряд, но их передний мозг уже слегка покрывает промежуточный; коры мозга у них еще нет, но в стенках переднего мозга намечается гистологическая дифференцировка. Хорошо развиты обонятельные и зрительные доли мозга (Табл. 2.1; 3). Однако экстирпация полушарий мозга не нарушает выполнение таких важных биологических функций амфибий, как плавание, схватывание живой добычи и даже спаривание.
У амфибий во взрослом состоянии утрачивается так хорошо представленный у рыб орган боковой линии, воспринимающий колебания, вибрации воды; органами осязания амфибий являются не только кожа, но и специальные осязательные бородавки. Хотя вся кожа амфибий способна воспринимать химические раздражения, но их органы вкуса сосредоточиваются во рту.
Саламандры способны различать 2 %-ный раствор сахара и 0,03 %-ный раствор хинина. Язык саламандр чувствителен к кислотам и солям. У наземных амфибий химическая чувствительность ниже, чем у водных. У амфибий в коже найдены раздельные тепловые и холодовые точки, а также точки, способные воспринимать прикосновение и боль. Это указывает на их чувствительность к перемене температуры, механическим и болевым воздействиям. Известно, что температурный фактор в жизни амфибий играет большую роль, так как при снижении t° до +7, +8° они впадают в состояние холодового оцепенения, зарываются в мох, в ил, под корни деревьев, где и перезимовывают до весны и потепления.
Органы обоняния амфибий, расположенные в передней части головы, более сложны, чем у рыб, и играют одновременно со зрением большую роль при отыскивании пищи. При опускании в аквариум мешочка с червяками тритоны тотчас чуют их и начинают искать, наклонив голову.
Гораздо сложнее устроен и орган слуха амфибий; у них значительно более, чем у рыб, развита способность издавания сильных звуков (кваканья), особенно в период размножения при взаимном нахождении разных полов. В экспериментальной ситуации обнаружилось, что на звук хвостатые амфибии не реагируют, бесхвостые амфибии (лягушки) реагируют весьма ограниченно: они, например, никак не реагируют на такие звуки, как хлопанье в ладоши, свист, стрельба; при звуке в 50—1000 колебаний в секунду у них изменяется кривая дыхания, они настораживаются, но биологически значимые для них звуки, например, шлепанье по воде и искусственное кваканье вызывают ответные реакции лягушек — первое побуждает их к прыганью в воду, второе — к ответному кваканью[75].
Глаза амфибий приспособлены к видению на более далекое расстояние, чем глаза рыб, они дают более отчетливую картину окружающего. Так же, как и рыбы, амфибии воспринимают движение предметов, что связано с питанием некоторых видов амфибий живой движущейся добычей. Лягушка может умереть с голоду, но не схватит неподвижно сидящую близ нее муху; с другой стороны, она будет стараться захватить движущийся даже несъедобный предмет.
При осуществлении акта питания у лягушки происходит координация движений глаз и подвижного, выбрасываемого наружу липкого языка, обеспечивающего прилипание жертвы, которую лягушка втягивает в рот и заглатывает, обычно закрыв при этом глаза. При нарочитом прикреплении движущейся добычи, лягушка производит выбрасывание языка, а вслед затем осуществляет мнимое заглатывание жертвы. Древесные лягушки, заметив жертву, сами прыгают к добыче, у этих лягушек при отыскивании пищи обоняние играет ведущую роль, так как их добыча неподвижна.
О различении амфибиями формы и величины предметов сведений нет. Амфибии способны к различению цвета, но главным образом в области коротковолновой части спектра, они чувствительны к ультрафиолетовым лучам и особенно к фиолетовому цвету.
Координирование деятельности зрительных рецепторов с двигательной активностью амфибий явственно обнаруживается не только при акте питания, но и в поведении, связанном с инстинктом самосохранения. Инстинкт самосохранения, самозащиты у амфибий проявляется весьма своеобразно.
Одни амфибии в случае опасности раздувают легкие, вместе с тем раздувая все тело. Некоторые лягушки (рода Вотbiпае) в случае приближения опасных для них змей и птиц принимают позы «угрозы», т. е. закидывают кверху голову и поднимают ярко окрашенные конечности; в подобных случаях саламандры резко машут хвостом, обильно выделяя жидкость; некоторые крупные амфибии ведут себя более агрессивно, т. е. раскрывают рот, издавая скрипучий звук, и кусаются.
Чрезвычайно многообразно и порой весьма сложно инстинктивное поведение амфибий, относящееся к сохранению потомства. Подобно тому, как мы наблюдаем это в других группах животных, у разных видов амфибий, в зависимости от условий их жизни, различается степень сложности проявления этого инстинкта. Иногда наблюдается полное безучастие родителей к своему потомству, т. е. оставление оплодотворенной икры без надзора; иногда родительский инстинкт проявляется более сложно и выражается в осуществлении серии последовательно выполняемых действий, направленных на устранение существующих неблагоприятных условий при созревании молоди.
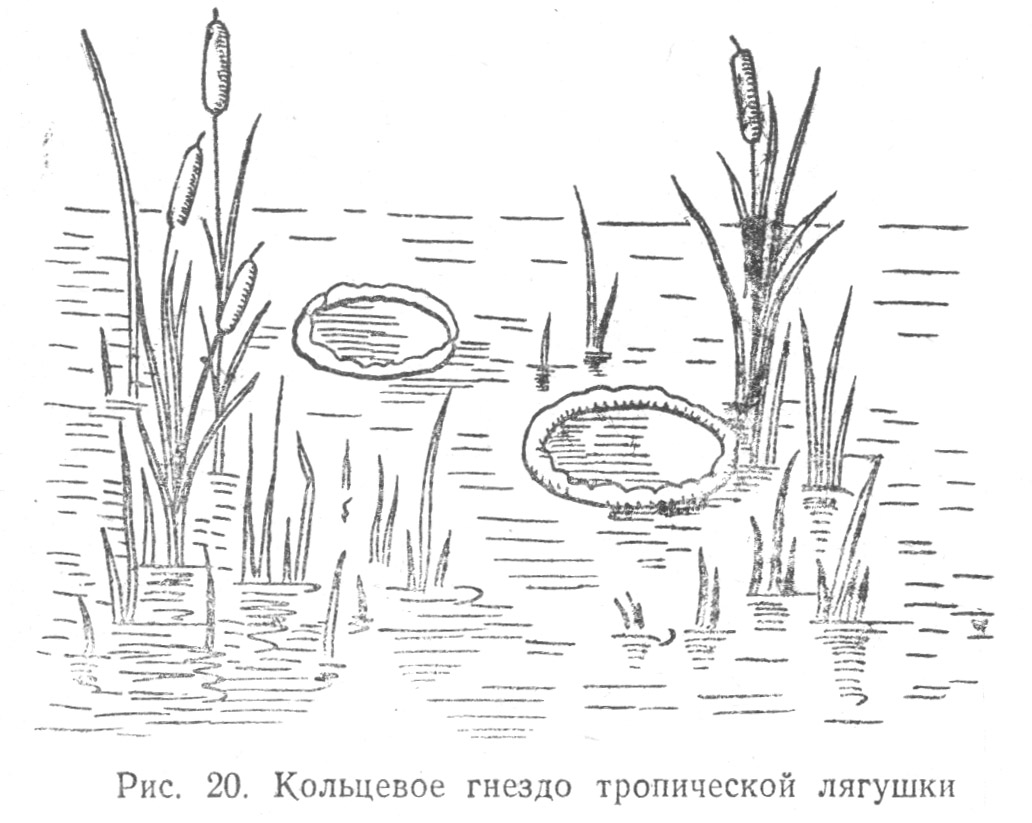
При быстром созревании икры, когда ее развитие продолжается не более суток, у амфибий за яйцами нет никакого ухода, но в тех случаях, когда появляется опасность утраты яиц из-за их поедания врагами, пересыхания, уноса водой, в поведении родителей возникает ряд приспособлений, обеспечивающих благополучное развитие яиц. Иногда яйца помещаются в надежное место для благоприятного созревания, иногда родители носят яйца с собой на ногах, на спине — в имеющихся на теле особых сумках или ячеях и даже во рту. Более того, некоторые амфибии (Hyla faber) даже устраивают специальные и сложные убежища, выступающие над водой в виде кольцевидно сооруженных гнезд (Табл. 2.3) из ила, принесенного в лапках, уложенного по краям гнезда и приглаженного ногами; ряд амфибий (Phyllomedusa hypohondralis) устраивает искусственные резервуары из листьев для откладывания в них икринок. У этой амфибии самка забирается на стебли прибрежных растений, сближает задними ногами края листьев, откладывая в получившуюся трубку икру, студенистая масса которой склеивает края листа.
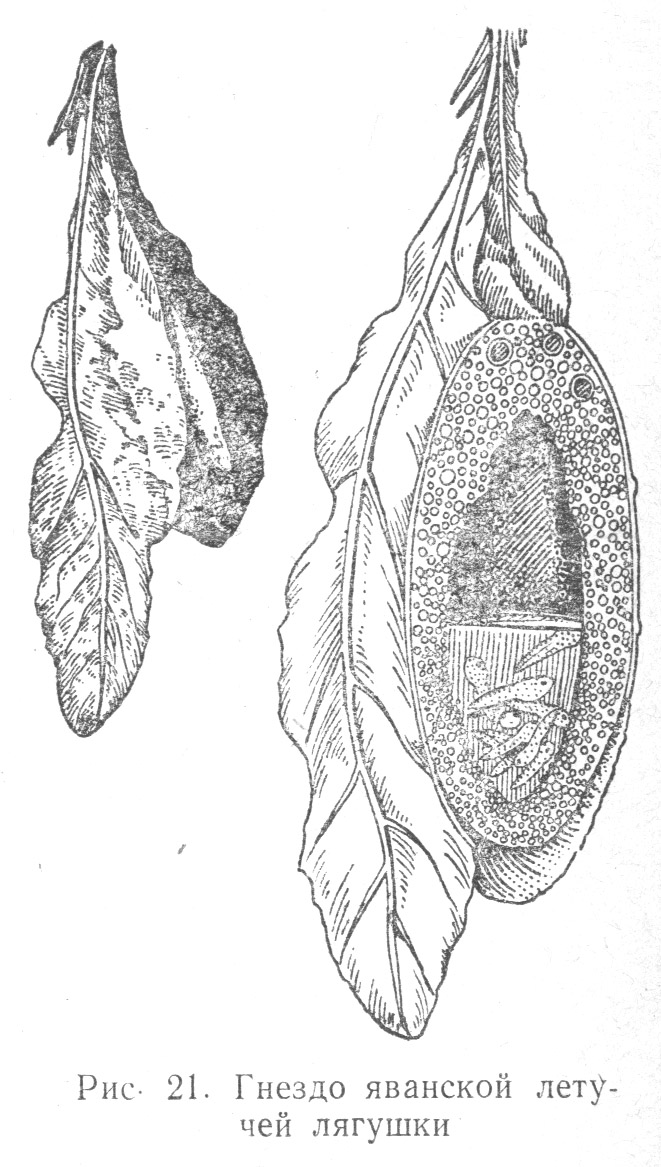
Яванская летучая лягушка (Rhacophorus Polypedotes rheinwaldtii) приклеивает к листьям пенистую массу, в которую погружает яйца (Табл. 2.4). Все эти сложнейшие формы поведения амфибий выработались и эволюционировали под влиянием внешней среды, закрепляясь в формах, наиболее выгодных для процветания вида.
Таким образом, у амфибий родители устанавливают определенные конкретные связи между биологически значимыми для них объектами (оплодотворенными яйцами) и собой, частями своего тела, благодаря наличию специальных приспособлений на нем для прикрепления, вмещения и сокрытия яиц. Они используют предметы окружающей среды, дифференцируя материал, пригодный для стройки (листья, ил, пенистую массу), они выделяют этот материал из окружающей среды, производят над ним определенные, не только обрабатывающие манипуляции, но и переносят его и конструктивно связывают отдельные части материала, например, делая круговые ограды из ила, приглаживая внутренние стенки кольцевого гнезда, сближая, склеивая края листьев[76].
Все эти действия могут осуществляться лишь при условии дифференцировки животными предметов, т. е. практического анализа окружающей среды и практического синтезирования предметов, включенных в цепь в основном стереотипного инстинктивного проявления поведения животных, выражающегося в уходе за потомством. Этот практический анализ и синтез, конечно, осуществляется на базе довольно сложной аналитико-синтетической деятельности их мозга.
Земноводные не превосходят рыб в скорости образования условных рефлексов при различного рода «обучении», хотя их передний мозг развит значительнее, чем у рыб, и, быть может, мы могли бы ожидать лучшей их «обучаемости».
Доказана способность амфибий к установлению условнорефлекторных связей. Но эти их ассоциативные способности весьма ограничены.
У наземных амфибий, особенно у жаб, быстро вырабатываются условные пищевые рефлексы: после 4—7 проб неприятных на вкус мохнатых гусениц жаба отказывалась их брать. Точно так же после немногих опытов жабы переставали брать движущиеся кусочки бумаги. Иногда обычное пищевое поведение амфибий затормаживается после одной пробы рыжего муравья или червя, обмакнутого в какой-либо ядовитый химический раствор. У лягушки легко выработался отрицательный условный рефлекс не схватывать насекомое, после того как это схватывание сопровождалось электрическим ударом (опыты Шеффера)[77].
С другой стороны, лягушки и тритоны длительно не могли приобрести способность затормозить схватывание видимой, но недосягаемой пищи, например, находящейся за стеклом, или заключенной в целлулоидный стакан. Они продолжали попытки схватить подобную пищу даже при введении отрицательных раздражителей, например, в том случае, когда кожа их челюстей сильно повреждалась острыми иглами, окружавшими приманку.
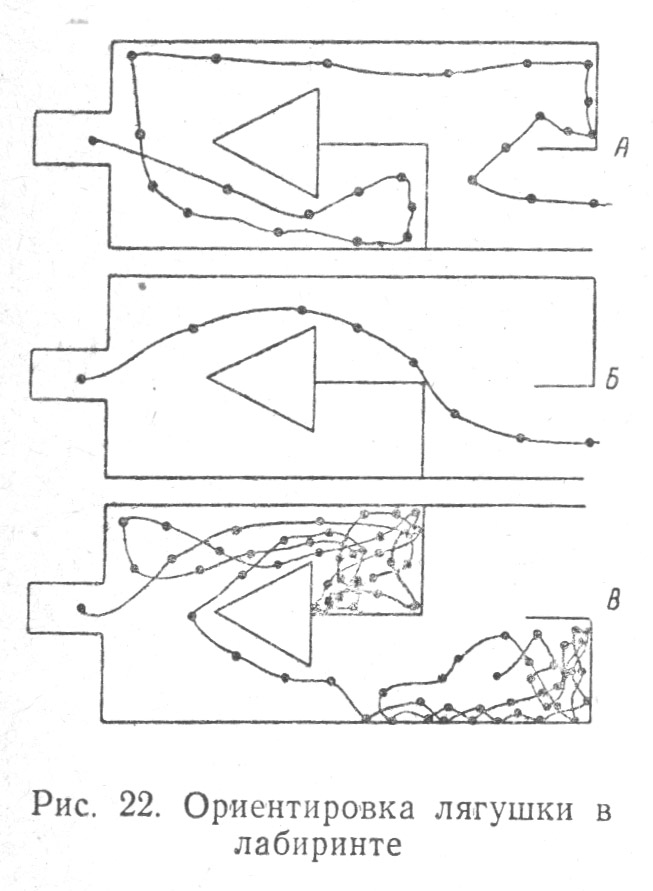
Помещенная в лабиринт лягушка через 120 опытов научилась выбираться к выходу наиболее кратким путем, но при перестановке лабиринта лягушка действовала согласно уже образовавшемуся навыку и не могла быстро переучиться, чтобы выбраться из тупиков к выходу (Табл. 2.5).
При окрашивании стенок лабиринта — одной в белый, другой в красный цвет — научение лягушки в лабиринте пошло быстрее, но при изменении окраски на обратную, лягушка не могла ориентироваться по цвету положительного стимула, приуроченного к определенному направлению коридора лабиринта и сбивалась в решении. При ориентировке животного в лабиринте обнаружилось, что зрительные стимулы имеют подсобное значение в отношении осязательно-кинестетических.
Испытанные в отношении ориентировки в лабиринте тритоны находили прямой путь к сосуду, содержащему пищу (опыты Пьерона). Бойтендайк добился того, что жабы научились обходить стеклянную пластинку, чтобы получить находящуюся за ней пищу[78].
Жабы оказались более способными, чем лягушки, к использованию обходных путей, ведущих к достижению пищи, помещенной за стеклянной пластинкой; лягушка не могла осуществить этот обход, так как непрерывно смотрела на пищу, жаба же временно отходила от видимой пищи с тем, чтобы обойти препятствие.
На основании возвращения сухопутных амфибий, жаб к своим норам, а лягушек к водоемам после удаления от них на значительное расстояние установлено наличие топографической, памяти у амфибий. Некоторые экспериментаторы (Баяндуров, Беритов) приходят к заключению, что образовавшиеся условные связи лягушек отличаются инертностью, непластичностью и лишают животное возможности переучивания при перемене значения положительного стимула на отрицательный и обратно[79].
Формы отражения окружающего мира у амфибий мало отличаются от форм отражения у рыб.
Отметим утрату ненужной для взрослых форм земноводных вибрационной чувствительности, столь полезной для типичных водных обитателей — рыб и, наоборот, улучшение более необходимой для жизни в воздушной среде зрительной и слуховой рецепции, весьма расширяющей анализ окружающего, участвующей в осуществлении таких жизненно важных актов, как например, питание, размножение.
Амфибии могут реагировать только на ограниченный круг сигнальных признаков, который для них остается постоянным, например, движение добычи или запах ее, вызывающие реакцию схватывания. Съедобный предмет, как таковой, не воспринимается лягушками как пища, если он неподвижен. Следовательно, в предмете выделяется лишь свойство, а не сам предмет: то подвижность, то запах предмета. У амфибий, несомненно, более сложен, чем у рыб, практический синтез раздражителей, обнаруживающийся как при акте питания, так и, в особенности, в многообразной конструктивной их деятельности при гнездостроении.
В акте питания мы наблюдаем у амфибий интеграцию зрительной и обонятельной рецепции с осязательной, вкусовой и двигательной.
В процессах сложнейшего конструирования гнезд, воспроизводимых при участии зрения, осязания, кинестезии, в актах выбора, переноса, обработки, размещения, скрепления различных материалов (с привлечением глины, пены, листьев и других предметов среды) мы имеем усложненную видовую, стереотипно протекающую деятельность амфибий, связанную с соответственно усложненной аналитико-синтетической деятельностью их мозга.
В неменьшей степени, чем у рыб, у земноводных доказано наличие способности к индивидуальному использованию опыта. При этом для установления условнорефлекторных связей сигнальными раздражителями являются главным образом химические и зрительные.
У амфибий наземных (жаб) в большей степени, чем у водных (лягушек), развиты тормозные процессы мозга.
Водные амфибии оказались малоспособными к перестройке поведения при изменении сигнального значения раздражителей — пространственных и цветовых.
[75] R. М. Yerkеs. The instincts, habits and reactions of the Frog. I. Associative processes of the green Frog. III. Auditory reactions of Frog. Harvard Psych. Studies, I, 1903, p. 627. R. M. Yerkes. The Sense of Hearing in Frogs, J. Compar neurol. and Physiol. V., XV, 1905, p. 297. См. также Д. H. Кашкаров. Современные успехи зоопсихологии. Гиз., 1928, стр. 326 и 327.
[76] О биологии амфибий. См. обширную библиографию в книге А. Г. Банникова и М. Н. Денисова. «Очерки по биологии земноводных». Учпедгиз, 1956.
[77] A. A. Sсhаeffer. Habit formation in Frogs. J. Animal Behavior, v. I, № 5, 1911.
[78] С. J. Warden, Т. N. Jenkins, L. Н. Warner. Comparative Psychologye (Vertebrates). New-York, 1936, p. 134—135.
[79] Б. И. Баяндуров. Условные рефлексы у птиц. Изд-во «Красное знамя», Томск, 1937, стр. 105.