Расширение среды для осуществления передвижения, способность к полету в воздухе, присущая представителям более высокоорганизованных, по сравнению с рептилиями, организмов — именно птиц — связана с возникновением у них ряда приспособительных прогрессивных морфолого- анатомических признаков и с появлением более высокого уровня отражения, психики этих организмов. Птиц следует рассматривать как прогрессивную ветвь животных, происшедших от пресмыкающихся, приспособившихся к полету в воздухе.
Наличие передних конечностей, крыльев с летательной поверхностью из перьев, дает возможность передвижения в воздушном пространстве. Разнообразно строение задних конечностей птиц — ног, приспособленных к передвижению в разных средах обитания: в лесах, в открытых пространствах (луга, степи и пустыни), в водной среде (озера и болота). Челюсти видоизменены в роговой клюв, имеющий весьма разнообразное строение у разных отрядов птиц и превосходно приспособленный к схватыванию, удерживанию, убиванию различного рода живой и захватыванию растительной пищи (ягод, семян растений). Все это отразилось на поведении птиц, связанном с основной формой их жизнедеятельности, способами добывания весьма разнообразной пищи.
Естественно, что широта распространения птиц по всему земному шару, появившиеся широкие возможности соприкосновения и связи птиц с разнообразнейшими предметами, встречаемыми в весьма различных по характеру средах их обитания, и обусловили большое развитие их нервной системы и органов чувств, их ориентировочной деятельности.
У птиц весьма велики большие полушария переднего мозга, а в них — полосатые тела (Табл. 2.1; 5 ); в связи с острым зрением птиц особое развитие получают зрительные доли их среднего мозга и перекрещивающиеся зрительные нервы. Перерезка зрительных нервов у голубя с одной стороны вызывает слепоту голубя на глаз другой стороны. Птица с удаленными полушариями неспособна узнавать знакомые предметы (например, птица-самка перестает узнавать своих птенцов).
При сравнительно большом объеме полушарий переднего, мозга для птиц характерна малая величина обонятельных долей (и обусловленное этим слабое развитие обоняния), большее развитие мозжечка, помогающего координации их движений и сохранению равновесия птиц при полетах.
Необычайное развитие получили большие по объему глаза птиц, способные к аккомодации, и не только к монокулярному, но у некоторых птиц и к бинокулярному зрению. Острота зрения некоторых хищных птиц превосходит таковую человека. Хищная птица — сокол сапсан ( Falco peregrinus) видит на земле небольших птичек величиной с горлиц с расстояния свыше 1077 м[84].
Экспериментально доказано, что птицы способны различать цвета так же хорошо, как человек, но оттенки цвета они видят хуже, чем человек. Они различают формы предметов (круг, треугольник, прямоугольник), рисунки (горизонтальные полосы), величины, изображения (в виде разного количества пятен), причем по степени различения они лучше всего дифференцируют величину, хуже степень освещенности и цвет и еще хуже форму предметов. Доказано, что птицы способны различать направление движения предметов.
Столь высоко развитое зрение имеет для птиц огромное значение в связи с их ориентацией в беспредельном воздушном пространстве при полетах, при опознавании ими вех на пути возвращения к гнездам, при обнаружении и ловле живой, тоже летающей добычи, при сезонных перелетах птиц за тысячи километров от мест их обитания, в связи с наступлением зимы и неблагоприятными условиями добывания корма.
При испытании слухового анализатора птиц (голубей) оказалось, что они различали свист, звук рожка, звонок, стук метронома; они дифференцировали силу, высоту и тембр звуков. Верхняя граница слышимости птицами звуков лежала между 11 000—12 000 колебаний в секунду. Птицы дифференцировали высокие тона хуже, чем низкие. В пределах третьей и четвертой октав голуби были способны различать полутона, в шестой октаве они различали лишь терции[85].
Слух птиц по отношению к биологически значимым раздражителям значительно более развит, чем у рептилий; птицы реагировали на еле слышимые человеком шумы. Тонкость слуха птиц подтверждается их способностью к пению, тончайшему модулированию своего голоса, различающемуся у разных видов птиц по высоте, силе звука и ритму, и замечательной способности птиц к подражанию не только пению и звукам птиц других видов, но и голосам других животных и даже голосу человека.
Птица-пересмешник может подражать звукам дятла, издаваемым им ударами клюва по дерезу, мяуканью кошки, воркованью голубя, блеянию ягненка, карканью ворона. Некоторые вороновые птицы (во́роны, воро́ны, галки), а также скворцы и особенно попугаи превосходно имитируют звуки вздоха, кашля, чмоканья губами, щелканья пальцами, свиста и пения и даже членораздельной речи человека, точно воспроизводя интонацию и тембр голоса.
Конечно, такое развитие слуха и голосовых способностей имеет для птиц большое биологическое значение, например, при обнаружении птицами издали живой добычи, при взаимных поисках самцов и самок в период размножения, при уходе за молодью. Известны призывные крики птиц, крики тревоги, угрозы и другие, связанные с выражением эмоциональных состояний птиц и косвенно служащие для особей своего вида сигналами к проявлению того или другого характера поведения (например, при подходе к партнеру для спаривания, избегании опасности, при отпугивании врагов — т. е. при пассивно- или активно-оборонительных реакциях птиц).
Все эти звуки, конечно, не являются направленными сигналами и выражают лишь определенные эмоциональные состояния птиц, но они воспринимаются сочленами группы как биологически значимые для них раздражители среды; вызывая приспособительные положительные или отрицательные реакции.
Менее развиты анатомически и менее значимы функционально для птиц обонятельный, вкусовой и осязательный рецепторы.
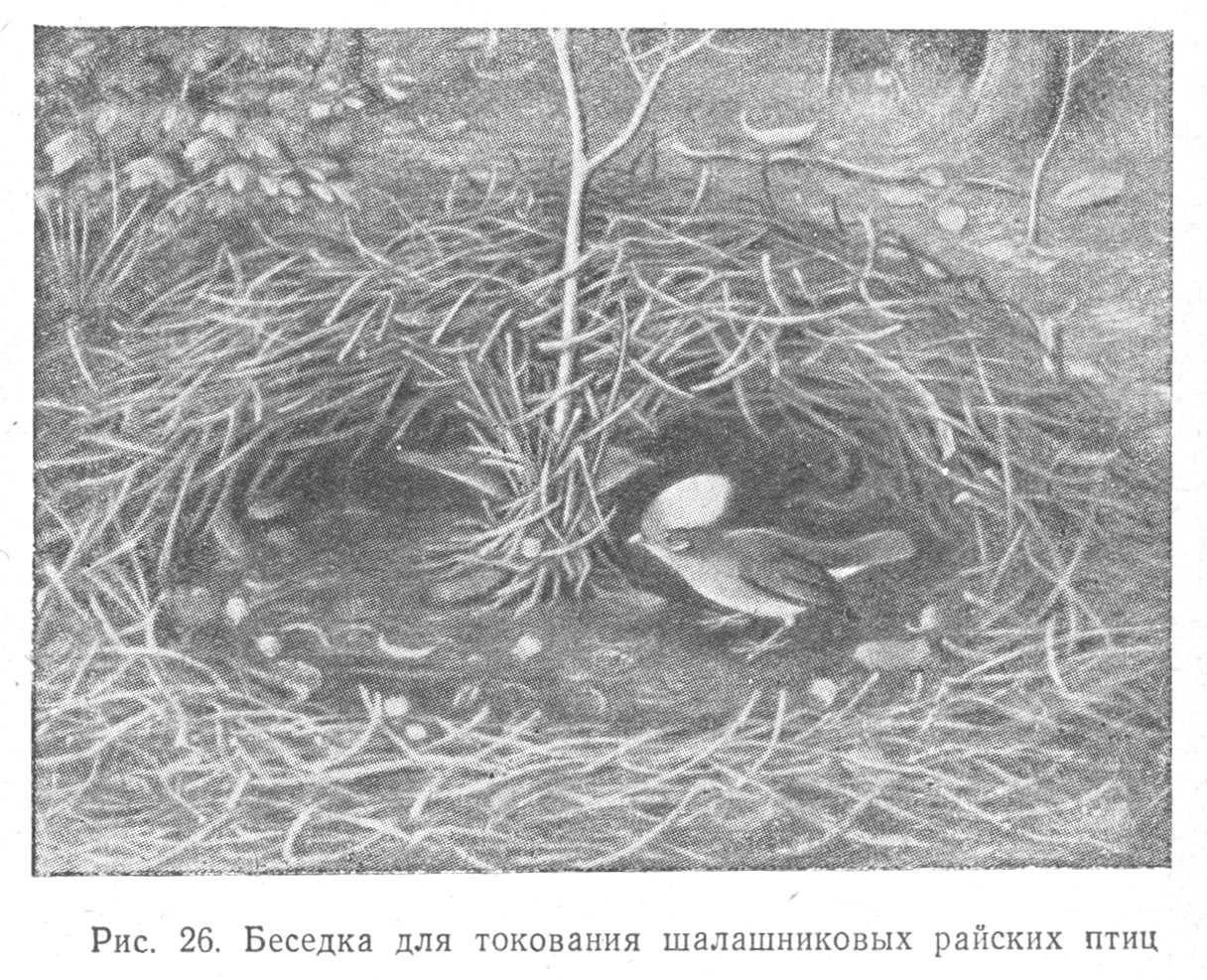
Принимая во внимание сложные формы приспособительного поведения птиц, относящиеся к питанию, к самозащите, к брачным играм (токам), сопровождаемым воспроизведением специальных поз, телодвижений (подобия «танцев»), постройкой своеобразных шалашей и беседок, например, у шалашниковых райских птиц (Табл. 2.9), наблюдая замечательное гнездостроение птиц, мы должны определенно признать, что аналитико-синтетическая деятельность мозга птиц находится на значительно более высоком уровне по сравнению с таковой у рептилий.
Действительно, мы замечаем у птиц чрезвычайное расширение диапазона раздражителей, вызывающих деятельность их зрительных и слуховых рецепторов и интеграцию их восприятий в сложных комплексах эффекторных реакций птиц. Этими раздражителями, выделяемыми птицами из внешней среды, являются и живые, и неживые объекты, находящиеся в самых различных средах обитания в сложном соотношении с окружающим.
Следует подчеркнуть, что добыча птиц чрезвычайно разнообразна, начиная от млекопитающих, являющихся нередко жертвой крупных хищных птиц, мелких птиц, рептилий, амфибий, рыб, насекомых, червей и кончая плодами и семенами разных растений.
Ловля хищниками добычи в воздухе, на лету, требует большой и точной координации зрения и кинестезии. Она нередко начинается, когда хищник находится еще на далеком расстоянии от местонахождения жертвы. Хищная птица, обозревая с высоты землю, находит притаившуюся живую добычу и не выпускает ее из поля своего зрения; при передвижении жертвы птица неотступно фиксирует ее глазами и координирует свою зрительную и двигательную деятельность, намечая путь полета и спуска, для овладения добычей.
Высматривание некоторыми птицами добычи с воздуха, подкарауливание ее появления из воды (у хищных птиц, питающихся рыбой, — например, чаек), вылавливание добычи в воде путем ныряния под водой также требует тончайшей зрительной дифференцировки и анализа окружающего.
У птиц, питающихся путем процеживания воды сквозь клюв-цедилку (у уток) или посредством обшаривания щелей под корой деревьев клювом и языком (у дятла), в акте питания принимает участие и осязательная рецепция.
У растительноядных птиц, употребляющих в пищу семена растений, практический анализ окружающих предметов путем зрительных дифференцировок направлен на различение съедобных для них растений, выделение частей этих растений (плодов, семян, ягод), на обработку этих выделенных частей до момента освоения съедобной части.
Особенно интересно, что этот практический анализ при выделении съедобных частей у разных видов птиц различен по отношению к одному и тому же виду растений. Например, плод вишни одни птицы (дрозды) поглощают целиком, другие (воробьи) ощипывают только мякоть плода, третьи (дубоносы) вылущивают и поглощают лишь косточку плода, бросая мякоть, как несъедобное. Эти особенности питания наследуются.
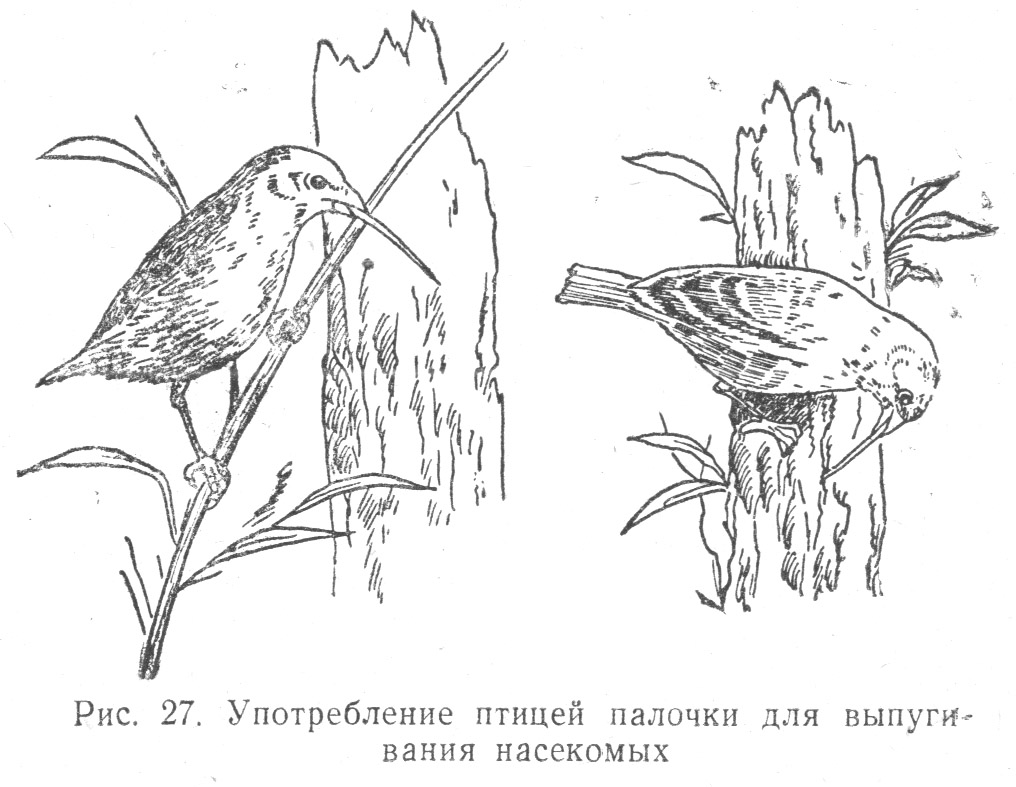
Отметим еще замечательную, носящую прирожденный характер способность одного вида птиц — именно дятлового древесного вьюрка (Camarhynchus pallidus) — использовать предмет (кактусовую иглу или веточку) для выпугивания из-под коры дерева насекомых, которыми эта птица питается. Наблюдатели передают, что птицы обычно отламывают веточку подходящей длины, отбрасывая слишком короткую или слишком длинную и заменяя ее соответственной[86] (Табл. 2.10).
Высокая степень зрительного анализа окружающего у птиц должна проявляться при поисках мест, подходящих для гнездования, в виде готовых убежищ (ямок, норок, дупел, расщелин скал), зарослей кустарника, крон высоких деревьев.
Тонкий, преимущественно зрительный практический анализ сопровождает выбор птицами подходящего гнездостроительного материала, специфического для разных видов птиц, — травы, мха, стеблей, прутьев, веток, шерсти, перьев, паутины, волос, глины, ила и т. д.
Следует подчеркнуть, что птицы, исследованные в условиях неволи методом естественного эксперимента выдающимся советским орнитологом Промптовым[87] , обнаружили большую пластичность в выборе гнездостроительного материала, отступая от стандарта этого выбора, и при наличии искусственного материала (например, марлевых бинтов), более пригодного для гнездостроения, использовали его, а не имеющийся поблизости естественный, но менее пригодный для гнездовития растительный материал.
Все это определенно указывало, что птицы адаптивна используют свой индивидуальный опыт, нарушая стереотип видового опыта, инстинкта, т. е. включают условно-рефлекторную деятельность. На значение индивидуального опыта и упражнения в процессе конструирования гнезда ярко указывает тот факт, что молодая птица обычно вьет гнездо хуже старой. Более того, у птиц, воспитанных изолированно от сообщества, никогда не наблюдавших процесса стройки, замечается полное смешение гнездостроительных актов, отсутствие их последовательного и преемственного развития, что указывает на огромную роль подражания при осуществлении некоторыми птицами гнездостроения (опыты Промптова).
Сравнительно высокий уровень синтеза раздражителей, получаемых птицами, обнаруживается в их способности к установлению связей между предметами в актах конструктивной деятельности, относящейся к постройке замечательных шалашей для токования и искусных гнезд для выведения птенцов.
Шалашниковые райские птицы сооружают близ ствола дерева своеобразные круглые высокие арены — платформы из мха, по которым они бегают кругами во время токования. Другие шалашниковые птицы конструируют коридоры из прутьев, у входа в которые располагают гладкие камешки, белые раковины, цветы, заменяя их свежесорванными при увядании (Табл. 2.9).
Как говорит известный советский эколог Д. Н. Кашкаров, у птиц «эволюция гнездостроения шла в разных систематических группах по разным путям в связи с особенностями других приспособлений: к средам обитания, к пище и в связи с особенностями местообитаний»[88].
Вследствие появления у птиц теплокровности, перяного покрова возникло высиживание яиц и выкармливание птенцов, что обеспечивало не только сохранение яиц, но позднее и выращивание птенцов и уход родителей за птенцами.
По развитию птенцов птиц разделяют на две биологические группы — выводковых и птенцовых.
У выводковых птиц (например, куриных, уток и др.) вылупившиеся птенцы в первые же сутки по выходе из яйца покидают гнездо и могут самостоятельно клевать, следуя за матерью.
У птенцовых птиц (голубиных, воробьиных, хищных) птенцы, вылупившиеся из яйца, довольно беспомощны и долго остаются в гнезде, получая пищу от родителей. Именно у птенцовых гнезда, устраиваемые родителями, особенно искусны, так как служат не только для насиживания яиц, но и для выращивания молодняка в период его выкармливания (Табл. 2.11, Табл. 2.12).
И у птиц легко можно проследить эволюцию инстинкта сохранения потомства и ухода за ним.
Некоторые птицы, гнездящиеся на прибрежных скалах, откладывают свои яйца просто на землю (гагарка — Alcatorda), между гальками (крачки, чайки), причем в последнем случае яйца имеют настолько резко выраженную покровительственную окраску, что их трудно отличить от окружающих камешков. Другие птицы откладывают яйца в готовых убежищах: в углублениях земли, в ямках (кулики — Limicolae), в расщелинах скал, в дуплах (совы — Striges); внутри которых птицы постилают мягкий материал. Некоторые птицы проявляют большую активность в гнездостроении и сами вырывают норки в прибрежных пластах песка и земли (береговые ласточки — Riparia). Гнездостроение, связанное с долблением твердого субстрата, мы находим у дятлов ( Pici), выдалбливающих себе дупла на стволах деревьев. Далее можно обнаружить сооружение птицами на земле простейших подстилок под яйца из мха, травы, листьев (например, у гусей, кур — Anseres, Galli). Некоторые группы птиц, также гнездящихся на земле, уже вьют гнездо из трав и стеблей, например, коньки (Anthus).
Огромное большинство птиц помещают свитые гнезда на ветвях кустарников и деревьев, то в прикорневой зоне — на тонких ветках дерева, то на высоких вершинах, где складывают огромные гнезда из толстых сучьев, сверху застилаемых более мягким материалом (у дневных хищников — орлов, у врановых).
Для некоторых птиц характерна лепная гнездостроительная деятельность, связанная с обмазыванием глиной выдолбленного или сплетенного гнезда (у дроздов — Turdus). Фламинго воздвигают себе из ила возвышения до 1/2 м высотой, на верхушке которых делают углубление для помещения яиц и птенцов[89] (Табл. 2.11). Лепные гнезда, построенные из растений и склеенные слюной птиц, мы находим у ласточек (Hirunda).
В группе воробьиных птиц (например, у зяблика) можно найти помещаемые на ветвях деревьев чрезвычайно искусно построенные гнезда, сплетенные из различных материалов: толстые стенки гнезда сделаны из мха, тонких корней и прутиков; внутри гнездо выстлано волосом и перьями, а снаружи облицовано лишайниками и берестой под тон того ствола дерева, на котором гнездо находится, что хорошо скрывает гнездо на окружающем фоне.
Особенно искусно бывают сделаны гнезда камышевок, имеющие форму глубоких цилиндрических вместилищ, висящих среди вертикальных стеблей камышей, гнезда тропических ткачиков, сшитые из сближенных и скрепленных между собой листьев, и гнезда синицы-ремеза в виде варежки (Табл. 2.12) — из мягкого растительного материала.

Самка птицы-носорога откладывает яйца в дупла, где самец замуровывает ее глиной на все время высиживания птенцов; он кормит ее, освобождая из гнезда лишь ко времени вылупления птенцов.
Иногда при постройке гнезд сочетается роющая и конструктивная деятельность птиц.
Замечательны гнезда сорных кур (Megapodidae ), зарывающих свои яйца внутрь огромных куч (высотой до 110 см) растительных веществ, где через 45 дней, под действием развивающегося от гниения тепла, выводятся птенцы. Молодая птичка разбивает клювом верхний тупой конец яйца, вылупляется и с помощью крыльев выбирается наружу, кверху. Как только она достигает поверхности холма, она зевает, отряхивается и бежит прямо в соседние кусты. Но если такого птенца вторично зарыть в кучу, то он оттуда не может выбраться и погибает.

Таким образом, конструктивная деятельность птиц достигает большой сложности в актах гнездостроения, начинаясь от более Примитивной стадии устройства простой подстилки для яиц из травы, листьев, мха, располагаемых на земле, и до более сложного сооружения, когда гнездовое вместилище конструируется птицей путем свивания, сплетения (стеблей, мха, волокон растений, водорослей), сшивания из листьев, склеивания из грязи, глины, слепления затвердевающей слюной.
Из этих разнообразнейших материалов птицы устраивают разной сложности и разной формы гнезда в виде лоточка, корзинки, шара, реторты, варежки, чулка и т. д. (см. Табл. 2.12). Гнезда некоторых видов птиц облепляются сверху дополнительным материалом — листочками, растительными чешуйками, мхом, хорошо скрывающими гнездо на окружающем его фоне.
Анализ гнездостроительной активности птиц А. Н. Промптовым привел его к заключению, что то, что называют инстинктивным поведением птиц, есть равнодействующая сложнейшего сочетания прирожденных безусловнорефлекторных и условнорефлекторных реакций.
«Основная и характернейшая черта этого поведения — направленная активность — как раз и иллюстрирует это положение, так как она обусловливается течением и сменой различных биологически-адаптированных доминант, функционально объединяющих разные „этажи“ нервной системы и тесно связанных также с гуморальной динамикой организма»[90].
«Целесообразность же инстинктивного поведения, — согласно Промптову, — имеет глубоко эволюционные корни, так как по сути дела зависит от функционирования врожденных механизмов, отобранных и закрепленных длительным процессом естественного отбора. Взаимодействие этих, филогенетически сложившихся (прирожденных) компонентов поведения с онтогенетически и индивидуально развивающимися компонентами, обусловливает степень приспособления каждого вида, определяет формы его реагирования на меняющиеся условия среды...».
«В таком дарвиновском смысле, понимаем мы, — пишет А. Н. Промптов, — эволюцию инстинктов, которые, конечно, меняются, затухают, исчезают совсем или рождаются вновь, в неразрывной связи с анатомо- морфологической эволюцией вида, текущей в характерной для него и также эволюционирующей внешней среде»[91].
Но, отмечая высокое развитие конструктивной деятельности птиц, мы должны подчеркнуть, что эти их способности проявляются при определенных нормальных условиях жизнедеятельности. Отклонение от нормы вызывает нецелесообразные действия птиц, свидетельствующие о ситуационной связанности их восприятий, в общем о стереотипности их сложной инстинктивной деятельности. Например, если у чаек, гнездящихся на земле среди прибрежных камней, переложить яйца из ямки, где они лежали, в другое место, то птицы продолжают насиживать пустую ямку, хотя яйца лежат поодаль, но близ нее; если в той же гнездовой ямке яйца заменить камнями или другими округлыми деревянными предметами, птица-мать этого не замечает и продолжает насиживание[92].
Аналогичные опыты были проведены Б. И. Хотиным и Г. З. Рогинским над кайрами на птичьих базарах Новой Земли[93].
По сравнению с рептилиями у птиц в проявлении инстинкта охраны потомства появляются новые, гораздо более длительные и активные связи матери, а иногда и; обоих родителей с подрастающим поколением. Как уже было отмечено, птенцовые продолжительно, выкармливают детенышей, приучают их к полету и ловле живой добычи (у хищных птиц) посредством бросания родителями в воздух пойманной живой жертвы перед птенцами; таким образом и предоставляется возможность поимки добычи молодыми птицами. В этом случае имеет место установление у птиц сложных связей уже не только между особями своего вида и своим потомством, но и межвидовых отношений с другого вида животными.
Исследования А. Н. Промптова, произведенные в Колтушской лаборатории (1947 г.), показали, что биологический контакт поколений играет очень большую роль в жизни птиц, в формировании их видового стереотипа поведения. Птенцы, находящиеся в непосредственной связи с выкармливающими их родителями, на основе, «свойственной всем птицам способности к подражанию... „осваивают“ характерные для вида кормовые стации, виды кормов...»[94].
И этого типа связи, хотя и осуществляются в основном в русле видовой инстинктивной деятельности, но, конечно, у каждой особи в процессе ее развития они образуются на основе индивидуального опыта и довольно сложной аналитико-синтетической деятельности ее мозга.
В последнее время, работая также в Колтушской лаборатории, Е. В. Лукина[95] , анализируя взаимодействие врожденных и приобретенных реакций птиц при выявлении изменяющегося воздействия среды обитания на формирование их пищедобывательной и гнездостроительной деятельности, пришла к интересным выводам. Она доказала, что под влиянием человека при перемене условий содержания птиц в различных ландшафтах можно формировать пищедобывательные реакции некоторых птиц, изменяя их стереотипный видовой способ питания на новый. Точно так же возможно заставить птиц изменять место гнездования и стереотипный выбор определенного гнездового материала. Означенное, направленное изменение инстинктов птиц можно широко использовать в практическом отношении, например, при необходимости переселения птиц в лесозащитные полосы.
В жизни птиц наблюдается интереснейшее явление — это весенние и осенние перелеты, т. е. регулярные массовые их переселения из одной области в другую.
С наступлением весны из жарких стран к нам в среднюю полосу СССР начинают прилетать различные птицы.
«Весенний перелет, — пишет А. Н. Промптов, — открывается появлением грачей. Затем прилетают полевые жаворонки, скворцы и вслед за этими первыми переселенцами стремительно набегает волна массового появления сразу многих видов перелетных птиц»[96].
В. Н. Бостанжогло в своей большой работе о птицах Арало-Каспийских степей (1911) так описывает валовой весенний пролет в устье р. Урала:
«Уже с половины марта и почти весь апрель (ст. ст.) горизонт ни на минуту не остается свободным. Трудно даже сказать, в какое время дня и ночи птицы летят меньше, так как стон стоит в воздухе во всякий час. Торопливо несутся развернутым фронтом стаи всевозможных пород уток, выплывая из-за горизонта длинными темными линиями и с тревожным свистом проносясь над устьями реки; тесными клубками валят стайки куликов и ржанок, перегоняя грациозных кроншнепов и суетливых веретенников; озабоченно пролетают одинокие хищнику; мерно и величественно плывут цапли. В приморской полосе в это время птица собирается в невероятном количестве. Подобно вокзалу большой узловой станции, все полно движения, сутолоки, криков, но нетребовательны пассажиры и дорого их время. Переведя дух и заморив на скорую руку червяка, стаи снимаются и летят далее, сменяясь все вновь прибывающими. Только немногие птицы делают более продолжительные остановки и не торопятся с окончанием своего воздушного путешествия. Во второй половине апреля в устьях Урала наступает уже сравнительная тишина, и местные птицы уже давно оберегают занятые участки...»[97].
С конца лета обычно у птиц начинается новое великое переселение. «Стаями и в одиночку, днем, ночью, утренними или вечерними зорями, каждый вид со свойственными ему особенностями, двигаются птицы на зимовки — снова в те места, откуда они прилетели несколько месяцев назад. Теперь эта армия переселенцев еще многочисленнее — она сильно пополнилась молодняком, который впервые в жизни совершает полет»[98].
«
Осенью от нас улетают миллионы птиц. Конечно, многие гибнут по пути
от бесчисленных опасностей, связанных с перелетом. За сотни и тысячи
километров от своих гнездований они летят до тропической Африки, Индии,
Китая и Индокитая или же задерживаются на побережьях Средиземного моря или
у южных берегов Каспия.
В этих местах находятся зимовки большинства птиц, населяющих летом Европу и
север Азии. На зимовках перелетные птицы находят себе подходящую пищу,
удобные условия жизни и держатся там до весны[99].
Большинство птиц, пролетая во время весеннего перелета тысячи километров, возвращается именно в те места, где они вывелись. Если осенний перелет птиц вызывается сезонными изменениями внешних условий: наступлением холодов и уменьшением кормов, то весенний прилет с мест зимовок вызывается в значительной степени внутренним физиологическим состоянием организма, состоянием, предшествующим периоду размножения птиц, связанного с определенными экологическими условиями — с поисками традиционной для вида гнездовой стации, — приводящими птиц в область старого гнездования, даже прошлогоднего гнезда.
Чем же руководствуются птицы в своих сезонных передвижениях, пролетая огромные пространства между двумя совершенно определенными географическими областями?
В этом отношении имеется много разных предположений. Одни ученые выдвигают значение физических факторов, вплоть до влияния земного магнетизма; некоторые ученые (Цион) на основании исследования ориентировки в пространстве почтовых голубей ссылаются на наличие у птиц специальной чувствительности к восприятию температурных изменений, интенсивности и направления ветра, ориентирующих птицу при полете. Ряд исследователей придает большое значение зрению птиц, их прекрасной зрительной памяти.
Другие авторы, занимавшиеся этим вопросом, подчеркивают связь зрительного и мускульного запоминания общей трассы перелета, или объединения восприятия нескольких органов при учете долготно-широтных координат при ориентировке птиц в пути. Пути перелета птиц являются определенными, складывавшимися тысячелетиями.
В перелетах птиц имеет громадное значение видовая инстинктивная традиция, передаваемая от поколения к поколению во время совместных странствований старых птиц с молодыми. Этот миграционный инстинкт является пережитком далекого прошлого птиц и благодаря ему древние зимовки, иногда, вследствие расселения вида, ставшие очень далекими, остаются все еще прочно связанными с его современным гнездовым ареалом.
Справедливо говорит А. Н. Промптов, что для разработки вопросов миграции необходим комплексный метод исследования на основе данных зоогеографии, экологии, физиологии и генетики. Это сложное явление не может быть правильно понято одним анализом составляющих его компонентов, изучаемых разными методами этих наук.
Переходя к особенностям индивидуально приобретенного, собственно условнорефлекторного поведения птиц, как оно обнаруживается в экспериментальной ситуации[100] , следует сказать, что самые различные зрительные раздражители могут быть для птиц сигнальными положительными раздражителями, например, свет, темнота, цвет, форма, величина фигур, количество пятен, направление движущихся фигур.
В своих экспериментальных исследованиях[101] зрительных дифференцировок у различных видов попугаев мы имели убедительные доказательства, что попугаи: ара, лори, какаду легко вырабатывают условные рефлексы на тот или другой цвет: красный, синий, зеленый, помещенный среди различных цветовых компонентов (Табл. 2.13).
В Колтушской лаборатории посредством применения «хватательной методики», в частности вызова пищедобывательного движения птиц (кур, голубей, ворон, галок, уток) путем дерганья клювом кольца или нажима клювом на рычаг, удалось выработать у птиц стойкие условные рефлексы и дифференцировку самых различных зрительных и слуховых раздражителей — зеленого и белого цвета, звука зуммера, метронома, звонка, — сочетаемых с подкреплением пищей. Следует отметить, что эти рефлексы сохранились после 5-месячного перерыва в работе с птицами[102].
При сравнении деятельности различных анализаторов птиц в процессе выработки условных рефлексов на дифференцировку раздражителей оказалось, что наилучшее дифференцирование давали зрительный и слуховой анализаторы[103] . Значительно хуже осуществлялась обонятельная дифференцировка различных запахов (одеколона, ментола, валерьяны); еще хуже действовал пространственный анализатор (дифференцировка направления вращения) и временной (образование условного рефлекса на получасовые промежутки).
У журавлей (Grus collarus) в зоологическом саду (в Галле) был выработан условный рефлекс приходить на свист к месту кормежки. Этот навык сохранился вопреки перерыву в подаче свиста в течение 21½ месяца.
В одном совхозе у гусей образовался условный рефлекс — целыми стаями прибегать с озера по звону колокола, извещавшего рабочих о наступлении часа обеденного перерыва. Куры выучились находить кратчайший путь в лабиринте, при прохождении которого надо было приобрести (выбирая один путь из двух) 23 различных условных рефлекса, составляющих сложную ассоциативную связь, обеспечивающую решение задачи. Но в более сложной обстановке куры, например, не могли найти обходный путь к пище, брошенной за решеткой, хотя требовалось только отойти от пищи и обойти решетку (и этот путь был им известен). Собака, а тем более обезьяна, легко решала подобную задачу (опыты В. Кёлера)[104].
Много опытов было поставлено с целью проверить способность различных видов птиц к ориентировке в лабиринте, к освоению и запоминанию ими кратчайшего пути выхода из лабиринта (опыты М. П. Садовниковой)[105].
На основании этих исследований установлено, что при ориентировке в лабиринте у снегиря вначале ведущую роль играет зрение, позднее, после освоения с лабиринтом, кинестезия. Изменение положения того же лабиринта — его поворот на 90°, 180°, 270° дезориентирует птицу, и она путается в прохождении; только в результате новых опытов она опять находит правильный и кратчайший путь выхода.

Испытанные по методу лабиринта голуби после прохождения одного лабиринта легче научились находить путь во втором лабиринте, разнящемся по конструкции от первого (опыты Роузе).
Голубей приучали открывать дверцу ящика с кормом, а воробьи были вытренированы открывать дверцу клетки с кормом путем потягивания ее за ниточку. Птицу-зеленушку приучили выбирать пищу из ящика, (находящегося среди четырех), причем она находила этот ящик и при перемене его местонахождения (опыты Бойтендайка).
Вороны оказались способными к образованию условного рефлекса на выбор определенного в ряду предмета. Так, из девяти открытых дверей они выбирали заданные: крайнюю правую или левую, но они не могли освоить выбор второй с краю левой двери[106].
Академик Бериташвили исследовал способность голубей к восприятию внешнего вида пищи, состоящей из большего (шести) или меньшего (трех) количества зерен. Опыты показали, что сначала голуби неизменно начинали клевать кучу, содержащую большее количество (шесть зерен); позднее они направлялись к кучке, содержащей меньшее количество (три зерна).
Когда же голубей движением руки стали отгонять от большей кучи, то через 250—260 опытов у них выработали условный рефлекс на клевание из меньшей кучки.
После 1001—1413 кормлений из кучки с тремя зернами голубь тянулся к этой кормушке, независимо от того, какое количество зерен находилось в соседней. Этот условный рефлекс закрепился настолько прочно, что вне клетки голубь клевал определенную кучку, состоящую из трех зерен; к другим кучкам он или не подходил, или подходил с большим промедлением[107].
Немецкий ученый О. Кёлер, исследовавший способность птиц к различению количества предметов, пришел к заключению, что птицы могут различать количество, определяя его «на глаз».
В его опытах голуби отличали пять зернышек от шести, когда обе группы располагались рядом. Более того, они приучились выклевывать из кучки только пять зерен, не трогая оставшиеся зерна. Волнистые попугаи выучивались различать количество шесть, и это количество было пределом их различения, а ворон, серый попугай и амазонский попугай могли различать до семи единиц.
Ворон, с которым О. Кёлер работал по нашей методике «выбора на образец», научился на показываемый ему образец-кружок с 3—4—5—6—7 черными точками открывать ту кормушку, на которой было изображено соответствующее количество точек[108].
Экспериментальное изучение способности к образованию условных рефлексов у птиц, в частности, у воробьев, дает возможности выявить некоторые их особенности при решении разного рода задач.
Такими особенностями являются: использование птицами своего индивидуального опыта, подражание одних индивидов опыту других при решении задач, наличие пробных, поисковых опытов, предваряющих правильное решение задачи, большая настойчивость в преодолении препятствий, способность переучиваться, приметливость к появлению новых элементов на фоне прежних, адаптация своих действий к меняющимся условиям, наличие хорошей памяти[109].
Поставленные в сравнительно-физиологическом плане опыты по изучению условных рефлексов у представителей позвоночных разных классов: рыб, черепах и птиц (галки) — при анализе подвижности корковых нервных процессов выявили возможность переделки у этих животных сигнального значения раздражителей, (именно зеленого и белого цвета), но повторная переделка лучше удалась у птиц, меньше у черепах и совсем не удалась у рыб[110] . Это происходило, по мнению автора, вследствие того, что птицы обладают большой тренируемостью подвижности нервных процессов по сравнению с черепахами и рыбами. У птиц удалось также выработать условные рефлексы второго порядка.
Баяндуров, сравнивая скорость образования условных рефлексов и их прочность у представителей пяти классов животных: рыб, амфибий, рептилий (черепах), а также птиц и млекопитающих (собак), подчеркивает, что чем выше в зоологической лестнице стоит животное, тем сильнее в его центральной нервной системе выражены тормозные процессы.
У собак они довольно значительны, гораздо интенсивнее, чем у птиц, у птиц значительнее, чем у рыб, амфибий или рептилий. И, наоборот, чем ниже на зоологической лестнице стоит животное, тем в его центральной нервной системе хуже протекают процессы внутреннего торможения условных рефлексов, а отсюда худшая дифференцировочная способность.
Согласно мнению Баяндурова, по способностям к образованию условных рефлексов птицы занимают среднее место между рыбами и млекопитающими и далеко отходят от амфибий и рептилий[111].
Обзор способности птиц к установлению условнорефлекторных связей в экспериментальной ситуации, выделение ими индифферентных раздражителей, сочетаемых с безусловным раздражителем, оценка точности дифференцировок раздражителей, адресованных к разным анализаторам, выявление прочности сохранения образованных навыков, их переделки при изменении значения раздражителей в новых ситуациях позволяет нам сделать заключение о характере аналитико-синтетических процессов у птиц.
Совокупность имеющихся экспериментальных данных обнаруживает сильно развитую способность птиц к аналитическому выделению самых различных по качеству раздражителей.
Обнаруживается способность птиц не только к выделению, но и тонкой дифференцировке многообразных раздражителей (зрительных, звуковых, в меньшей степени обонятельных, пространственных, временных) на фоне окружающей обстановки, что связано с участием тормозных процессов, которые у птиц явно более выражены, чем у рыб, амфибий и рептилий.
Совершенно несомненна довольно высокая способность птиц не только к синтезированию индифферентных раздражителей с биологически значимыми, но связывание их сенсорных дифференцировок с разнообразнейшей двигательной активностью птиц — движением клюва, ног, с актами дерганья, нажима, схватывания клювом, взятия и подачи клювом, с подбеганием к определенному раздражителю, прохождением лабиринта, подлетом и присаживанием на жердь и т. д.
Установленные синтетические связи птиц отличались большой прочностью. Выучка сохранялась у птиц при перерыве в работе от 5 до 21 месяца, в противоположность тому, что наблюдалось у амфибий (лягушек), у которых выработанные условные рефлексы приходилось образовывать заново в каждый последующий день экспериментов, и у рептилий (черепах), у которых угасание условного рефлекса шло медленнее, чем у амфибий, но скорее, чем у птиц.
Вопреки относительно большой устойчивости, установленные временные связи птиц отличались пластичностью и могли быть видоизменены, переделаны, а у испытанных в лаборатории рептилий (черепах), и в еще большей степени у рыб и амфибий такая переделка давалась с трудом.
С другой стороны, следует подчеркнуть, что в случае установления более сложных синтетических связей, последние, возникали у птиц в результате более или менее длительного поискового пути (проб и ошибок), предваряющего образование замыкательного процесса в мозгу птиц, обеспечивающего установление условной связи.
Эти установленные временные связи были действенны преимущественно в определенной ситуации, в которой они были получены, перенос их в измененные условия, сравнительно мало разнящиеся от прежних, содействовал более скорому образованию новых связей на базе существующих, например, при осложнении пути при ориентировке птицы в лабиринте.
Но при резкой перемене условий, например, повороте лабиринта на 90°—270°, требующей более радикальной перестройки образовавшихся двигательных навыков, птицы не могли сразу решить задачу. Птица снова нащупывала измененный путь, пробега, снова устанавливала связи для образования соответственного нового условного рефлекса. Прочно фиксированные у птиц, сравнительно мало пластичные условные связи в более сложных случаях препятствовали образованию новых условных связей. Этим, объясняется трудность доставания птицами корма, обнаруженная, например, у кур, при необходимости использования ими обходных путей. Как было упомянуто, курица, привыкшая брать видимый корм, непосредственно перед ней расположенный, настойчиво рвалась через сетку к этому корму, вопреки имеющемуся у нее (на основании прежнего опыта) знанию других путей выхода к корму; она явно уступала в отношении пластичности условных связей млекопитающим животным (собаке и обезьяне)[112].
Сравнивая уровень отражения у птиц с таковым ранее рассмотренных позвоночных животных (рыб, амфибий, рептилий), мы находим у них несомненное повышение и усложнение форм отражения.
Появление коры мозга и совершенствование высокоорганизованных органов чувств — зрения и слуха — определяет значительный прогресс анализа, особенно зрительного, как в отношении его необычайного обострения, так и возникновения цветного и предметного зрения. Аналогичное утончение и расширение диапазона рецепции имеется у птиц и в области слуха. В соответствии с этим наблюдается значительное расширение ориентировочной деятельности птиц. Аналитико-синтетическая деятельность мозга птиц приобретает более сложные формы вследствие возрастания многообразия связей организма со средой.
Птицы оказались способными к уточненному анализу отдельных свойств изолированных раздражителей окружающей среды как зрительных (света, тьмы, яркости, цвета, формы, величины, рисунка, движения и направления движения предметов), так и слуховых (стука, свиста, шума и высоты, силы, тембра, ритма звука).
Птицы оказались способными к синтезу получаемых раздражений в целостное восприятие при зрительном различении предметов, при подражательной репродукции тех же видов звуков, при воспроизведении ими голосов различных птиц и других животных и даже членораздельной человеческой речи.
Сложная аналитико-синтетическая деятельность птиц проявляется в актах питания, самосохранения, размножения, сохранения потомства, включающих участие огромного количества безусловных сигнальных раздражителей, вызывающих биологически целесообразные реакции птиц.
Напомним сложно осуществляемые акты питания, связанные, например, с высматриванием добычи, выбором съедобных плодов, акты конструирования (шалашей, беседок, платформ для толкования, гнезд для выведения птенцов), когда птица избирательно выделяет материал из сложного комплекса окружающей среды, когда она многообразными приемами осуществляет связи между самыми различными предметами, оформляя самые различные типы гнезд.
В отношении деятельности гнездостроения следует отметить, что наравне с натуральными, сигнальными раздражителями, строго определенными у разных видов птиц, оказываются действительными в силу генерализации раздражителей и другие, новые сигнальные стимулы, более или менее сходные с обычными. Так, например, птицы могли использовать для сплетения гнезд вместо стеблей растений случайно оказавшиеся поблизости предметы человеческого обихода — бинты — и даже предпочитали эти материалы естественным. Подобное изменение поведения указывает на возможность нарушения птицами видового динамического стереотипа при изменении внешних условий и на способность их к широкому использованию своего индивидуального опыта.
Прогрессивные особенности форм отражения птиц ярко сказываются в их способности к выработке навыков, т. е. условных рефлексов на самые различные индифферентные раздражители, сочетаемые с безусловными и ставшие сигнальными. Многообразие условных сигнальных раздражителей (зрительных, слуховых, обонятельных) свидетельствовало о том, что птицы были в состоянии осуществлять более дробный и углубленный по сравнению с рептилиями анализ новых свойств и качеств среды и более сложный новый синтез раздражителей на основе накопления своего индивидуального опыта и приспособительного его использования. Выработка у птиц условных рефлексов, связанная с различением ими многообразных зрительных и слуховых сигнальных признаков, так же как и при нахождении ими пути в лабиринте, указывает, что аналитико-синтетические процессы птиц состоят не только в дифференцировке ими отдельных свойств предметов, но и в целостном восприятии ими совокупности этих свойств, в восприятии комплексных раздражителей, и их соотношений в длительном, прочном их сохранении.
У птиц, по сравнению с рептилиями, сильнее выражены и тормозные процессы мозга и подвижность нервных процессов при переделке сигнального значения раздражителей. Синтез птицами индифферентных раздражителей в сложном интегрировании их в самых различных условнорефлекторных связях свидетельствует и об усложнении динамики нервных процессов птиц и более сложном взаимоотношении и взаимодействии механизмов, являющихся физиологической основой весьма углубившегося и уточнившегося отражения окружающего мира.
Но все же следует подчеркнуть сравнительно малую подвижность высшей нервной деятельности птиц, недостаточную пластичность их поведения при резкой перемене условий, отражающуюся в трудности переделки прежних условных связей и образования новых.
[84] Г. П. Дементьев. Руководство по зоологии, т. VI. Позвоночные. Птицы. Изд. АН СССР, 1940, стр. 207.
[85] Б. И. Баяндуров. Условные рефлексы у птиц. Изд-во «Красное знамя». Томск, 1937, стр. 103.
[86] Д. Лэк, Дарвиновы вьюрки. Изд. АН СССР, М., 1949, стр. 71.
[87] А. Н. Промптов. Физиологический анализ инстинкта гнездостроения у птиц. «Известия АН СССР» (серия биологическая), 1945, № 1, стр. 1—25.
[88] Д. Н. Кашкаров, В. В. Станчинский. Kуpc зоологии позвоночных животных, изд. 2 е, изд. АН СССР, 1940, стр. 641.
[89] D. Heinгоth. Merkwürdige Vogelnester. Wunder d. Natur, 1912, S. 74—80.
[90] А. Н. Промптов, Физиологический анализ инстинкта гнездостроения у птиц. «Известия АН СССР» (серия биологическая), 1945, № 1, стр. 24.
[91] А. Н. Промптов. Физиологический анализ инстинкта гнездостроения у птиц. «Известия АН СССР» (серия биологическая), 1945, № 1, стр. 24.
[92] Г. А. Скребицкий. Отношение чайки к гнезду, яйцам и птенцам. Сб. «Рефлексы, инстинкты и навыки», т. II, Госсоцэкгиз, 1936, стр. 139—178.
[93] Г. З. Рогинский. Сравнительно-психологическое изучение птичьих базаров Новой Земли. Сб. «Арктический институт», 1935.
[94] Е. В. Лукина. К вопросу о взаимодействии врожденных и приобретенных реакций в жизнедеятельности птиц. Труды Института физиологии им. И. П. Павлова, т. II, изд. АН СССР, 1953, стр. 343.
[95] См. там же, стр. 342.
[96] А. Н. Промптов. Сезонные миграции птиц, изд. АН СССР, 1941, стр. 5.
[97] Цитировано по книге А. Н. Промптова «Сезонные миграции птиц». Изд. АН СССР, 1941, стр. 6.
[98] Там же, стр. 7.
[99] Цитировано по книге А. Н. Промптова „Сезонные миграции птиц“. Изд. АН СССР, 1941, стр. 10—11.
[100] Б. И. Баяндуров. Условные рефлексы у птиц. Изд-во «Красное знамя», Томск, 1937.
[101] См. Н. Н. Ладыгина-Котс. Отчет о деятельности Зоопсихологической лаборатории при Государственном Дарвиновском музее. Огиз, М., 1921.
[102] А. В. Бару. Методика исследования двигательных и пищевых условных рефлексов у птиц и условное торможение у птиц. Труды Института физиологии им. И. П. Павлова, т. II, изд. АН СССР, 1951.
[103] См. опыты Б. И. Баяндурова. Условные рефлексы у птиц. Труды Томского мед. института, т. V, Томск, 1937, стр. 449—453.
[104] В. Кёлеp. Исследование интеллекта человекоподобных обезьян. Изд. Ком. академии, М. 1930, стр. 18—19 (пер. с немецк.).
[105] Д. Н. Кашкаров. Современные успехи зоопсихологии, М. — Л., 1928, стр. 95 и 343—344.
М. Sadownikowa. A study of the behavior of birds in the maze, J. Comparat. Psych, v. III, 1923, № 2.
[106] F. Hempelmann. Tierpsychologie. Akad. Verlagsgesellschaft. Lpz., 1926, S. 392.
[107] И. С. Беритов и М. Ахметели. О роли внешнего вида пищи в индивидуальном поведении голубей. Труды Института им. Бериташвили, Тбилиси, 1937, стр. 375—394.
[108] О. Koehler. Vom unbenannten Denken «Lebendiges Wissen». В. 99, Wiesbaden, 1953, S. 271—279.
[109] Д. H. Кашкapов. Современные успехи зоопсихологии, Гиз, М. — Л., 1928, стр. 342—343.
[110] См. О. В. Третьякова. О подвижности нервных процессов у рыб, черепах и птиц. Труды Института физиологии им. И. П. Павлова, т. II, М. — Л., 1951, стр. 418—432.
[111] Б. И. Баяндуров. Условные рефлексы у птиц. Изд-во «Красное знамя», Томск, 1937, стр. 106.
[112] В. Кёлер. Исследование интеллекта человекоподобных обезьян. Изд. Комакадемии, М., 1930, стр. 18—19 (пер. с немецк).